
Рефераты по рекламе
Рефераты по физике
Рефераты по философии
Рефераты по финансам
Рефераты по химии
Рефераты по хозяйственному праву
Рефераты по цифровым устройствам
Рефераты по экологическому праву
Рефераты по экономико-математическому моделированию
Рефераты по экономической географии
Рефераты по экономической теории
Рефераты по этике
Рефераты по юриспруденции
Рефераты по языковедению
Рефераты по юридическим наукам
Рефераты по истории
Рефераты по компьютерным наукам
Рефераты по медицинским наукам
Рефераты по финансовым наукам
Рефераты по управленческим наукам
Психология и педагогика
Промышленность производство
Биология и химия
Языкознание филология
Издательское дело и полиграфия
Рефераты по краеведению и этнографии
Рефераты по религии и мифологии
Рефераты по медицине
Рефераты по сексологии
Рефераты по информатике программированию
Краткое содержание произведений
Реферат: Строение, свойства и биологическая роль биотина и тиамина
Реферат: Строение, свойства и биологическая роль биотина и тиамина
Биотин
Исторические сведения
Более 60 лет назад Wilidiers показал, что для обеспечения жизнедеятельности Дрожжевых клеток в искусственную питательную среду необходимо внести какое-то органическое вещество (фактор роста). Этот фактор роста он предложил назвать «биосом» (от греч. «bios»—жизнь). Изучение природы «биоса» привело к заключению, что «биос» представляет собой комплекс факторов роста, отличающихся друг от друга по физико-химическим свойствам. Экстракты, содержащие «биос», при обработке уксуснокислым свинцом разделялись на две биологически активные фракции. Фракция, выпадавшая в осадок, была названа «биос I», а фракция, остававшаяся в растворе, получила название «биос II». В 1928 г. «биос I» выделен из чая и идентифицирован как инозит. В 30-х годах «биос II» подвергался тщательному изучению и был разделен на две фракции—«биос IIа» и «биос IIв»—путем адсорбции последней на животном угле.
Kogi предложил назвать «биос IIв» прото «биосом II» или биотином, а фракцию «биос IIа»—«биосом III». В 1935—1936 гг. Kogi и Tonnies впервые выделили кристаллический биотин из желтка яиц. Для этой цели они использовали 250 кг желтков яиц и получили 100 мг биотина с температурой плавления 148°. Позже было найдено, что некоторые виды Rhizobium требуют для своего роста какое-то органическое вещество, присутствующее в культурах Azotobacter в гидролизованных дрожжах и других естественных источниках. Это вещество получило название «коэнзим Р». Сравнительное изучение действия на рост Rhizobium «коэнзима Р» и кристаллического препарата биотина показало, что оба вещества обладают одинаковыми биологическими свойствами.
В 1931 г. Gyorgy (Gyorgy, 1954) в опытах на крысах обнаружил, что ряд естественных источников содержит вещество, предохраняющее животных от заболевания, вызываемого избытком белка яиц, и предложил назвать его витамином H. 1939 г. он получил препарат витамина Н из печени. В процессе изучения физико-химических свойств препаратов витамина Н и распространения его в естественных источниках возникла мысль об идентичности витамина Н и биотина. Экспериментальная проверка показала, что наиболее очищенные препараты витамина Н полностью заменяют коэнзим Р при испытании на культурах Rhizobium tritolii и биотин при испытании на дрожжах. С другой стороны, коэнзим Р или биотин полностью заменял витамин Н при испытании на животных. Таким образом, витамин Н и биотин полностью идентичны. Дальнейшие исследования дали возможность установить химическое строение биотина и осуществить его синтез.
2. Химические и физические свойства биотина
В 1941 г. du Vigneaud и сотрудники выделили из печени кристаллический препарат метилового эфира биотина, из которого путем омыления щелочью был получен свободный биотин. Эмпирическая формула его была определена как C10H16O3N2S. На основании изучения продуктов распада биотина эти авторы пришли к выводу, что структура витамина соответствует 2/-кeтo-3,4-имидaзoлидo-2-тeтpaгидpoтиофен-н-валериановой кислоте.
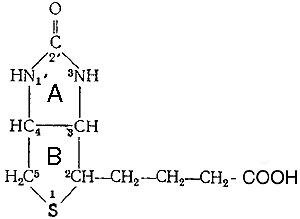
Молекула биотина состоит из имидазолового (А) и тиофенового (В) колец. Гетероцикл можно рассматривать как тиофеновое кольцо, связанное с уреидной группировкой. Приведенная структура биотина была подтверждена полным его химическим синтезом. В молекуле имеется три асимметрических атома углерода, что обусловливает существование 8 стереоизомеров.
Биотин образует игольчатые кристаллы с температурой плавления 232°. D поперечном сечении кристалл представляет собой ромб, острые углы которого равны 55°. Длина осей: а—5,25Ǻ, b—10,35Ǻ, с— 21,00Ǻ. Плотность кристаллов 1,41. Молекулярный вес биотина на основе химической формулы равен 214. Нa на основе рентгеноскопического анализа 245±6. При исследовании кристаллической структуры биотина установлено, что алифатическая цепь находится в цис-положении по отношению к уреидной циклической группировке.
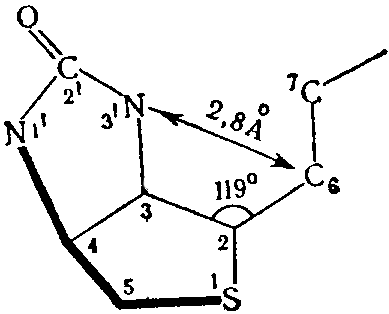
На основании данных, полученных методом рентгеноскопической кристаллографии Traub (1959) считает возможным образование внутримолекулярной водородной связи, которая возникает между кислородом карбонильной группы и одним из кислородных атомов карбоксильной группы в результате близкого расстояния между N-3’ и С-6 равного 2,4Ǻ; все остальные расстояния в молекуле составляют более 3,4Ǻ. Образование такой связи должно изменять распределение зарядов в уреидном кольце со смещением, кэтоэнольного. равновесия .к энолу, что: приводит к изменению химической реактивности N-1. Наличие водородной связи в известной мере определяет биологическую активность биотина и его производных. В 1965 г. установлена структура и относительная конфигурация каждого асимметрического центра биотина, а в 1966 г. абсолютная стереохимия витамина.
Биотин хорошо растворим в воде и спирте, трудно растворим в эфире, углеводородах парафинового ряда и несколько лучше в циклогексане, бензоле, галогенизированных углеводородах, спиртах и кетонах (ацетон). Биотин устойчив к действию ультрафиолетовых и рентгеновых лучей. Он разрушается под влиянием перекиси водорода, соляной кислоты, едких щелочей, формальдегида и сернистого газа. Он не изменяется под действием молекулярного кислорода, серной кислоты, гидро-
ксиламина. В ультрафиолетовых лучах для биотина не обнаружено специфического поглощения.
2.1 Аналоги и ингибиторы. Молекула биотина обладает большой специфичностью. Это подтверждается тем, что из 8 известных стереоизомеров только один—биотин—обладает биологической активностью. Удаление или замена каких либо атомов или групп атомов приводит к полной потере активности (дегидробиотин, гемоглобин, норбиотин). Только одна группа производных, полученных окислением атома серы (сульфобиотин и биотинсульфоксид) или заменой серы кислородом (оксибиотин) или двумя атомами водорода (дестиобиотин), проявляет биологическую активность. Сульфобиотин заменяет потребность в биотине у некоторых дрожжей но оказывается
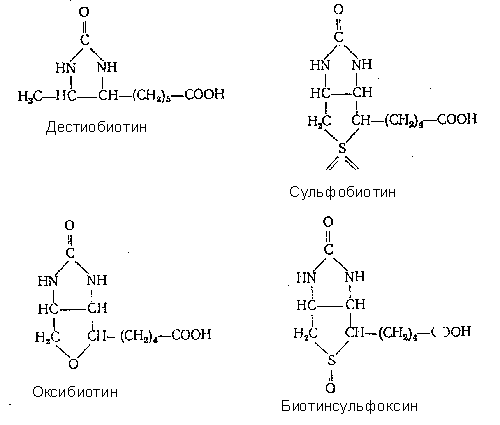
антагонистом витамина для L. casei, E. coli и Neurospora.
Известны аминокислотные производные биотина, среди которых наиболее изучен биоцитин, обладающий высокой активностью для многих микроорганизмов. Биоцитии выделен в кристаллическом виде из дрожжей. В 1951 г. расшифрована его структура. Оп представляет собой пептид биотина и лизина, а именно:
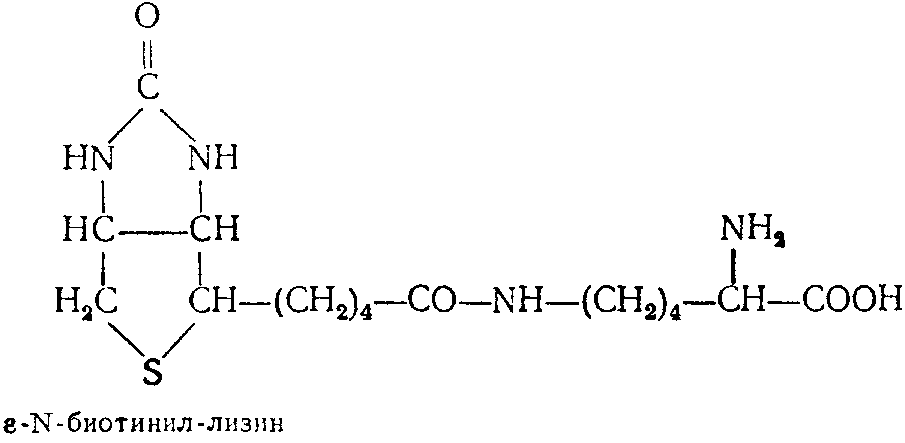
В 1952 г. осуществлен синтез биоцитина. Степень использования биоцитина разными микроорганизмами резко различается. Возможной причиной этого может быть наличие или отсутствие биоцитиназы, которая расщепляет биоцитин с освобождением свободного биотина.
По данным Traub (1959), биологическая активность биотина и его аналогов обусловлена внутримолекулярной связью, что позволяет объяснить причины наличия или отсутствия биологической активности для большинства изомеров и производных биотина. Так, образование водородной связи исключено у всех других оптических изомеров, кроме D-биотина, а также у производных с более длинной или укороченной боковой цепью, что приводит к полной потере биологической активности (гомобиотин, норбиотин). Неактивность гуанидиновых аналогов биотина, биотинола, оксибиотинола также объясняется отсутствием у них водородной связи. Некоторые производные биотина, которые образуются без нарушения водородной связи, сохраняют биологическую активность (дестиобиотин, оксибиотин, биоцитин и ряд аминокислотных производных биотина).
В настоящее время выяснена причина патологических изменений, возникающих при кормлении животных сырым яичным белком. В нем содержится авидин—белок, который специфически соединяется с биотином (введенным внутрь с пищевыми продуктами или синтезированным кишечными микроорганизмами) в неактивный комплекс и тем самым препятствует его всасыванию. Авидин содержится в яичном белке курицы, гуся, утки, индейки и лягушки. В 1942 г. он получен в кристаллическом виде и оказался глюкопротеидом с молекулярным весом 70000. Авидин стехиометричёски связывает эквимолярные количества биотина, образуя прочный комплекс, который не расщепляется ферментами пищеварительного тракта. Комплекс авидина с биотином термически устойчив и полностью диссоциирует только в автоклаве при 120° за 15 минут. С авидином соединяется DL-оксибиотин и некоторые другие аналоги биотина, но сродство авидина к биотину намного больше, чем к его производным. Изучение взаимодействия биотина и его производных с авидином показало необходимость уреидной группы в молекуле витамина, тогда; как карбоксильная группа и атом серы не являются необходимыми для образования комплекса. Авидин является универсальным ингибитором биотина. Его способность связывать биотин широко используется при изучении механизма участия этого витамина в процессах обмена веществ: торможение той или иной биохимической реакции авидином является существенным доводом в пользу возможности участия в ней биотина. Авидин применяется для получения экспериментальной биотиновой недостаточности у животных.
Распространение биотина в природе
Биотин широко распространен в природе. Он обнаружен у микроорганизмов. растении и животных. Содержание его определено в различных систематических группах животных: простейших, насекомых, рыб, земноводных, птиц, млекопитающих. Наиболее высокий уровень биотина обнаружен в личинках насекомых и наименьший — у пресмыкающихся. Рекордное количество (6,81 мкг/г) найдено в печени акулы. Содержание биотина в организме животных не зависит от принадлежности животного к определенной систематической группе. Анализ тканей показал большое различие в содержании биотина в органах одного и того же животного. Наиболее богаты витамином печень, почки, надпочечники; сердце и желудок содержат среднее, а мозговая ткань, легкие и скелетные мышцы—минимальное количество биотина.
Ниже приведено содержание биотина в различных продуктах животного и растительного происхождения.
Содержание биотина в пищевых продуктах (В. В. Филиппов, 1962)
|
Продукты животного происхождения |
Биотин в мкг на продукта |
Продукты животного 100 г происхождения |
Биотин в мкг на 100 г продукта |
|
Печень свиная |
250 |
Рисовые отруби |
46 |
|
» говяжья |
200 |
Рожь, цельное зерно |
46 |
|
Почки свиные |
180 |
Земляные орехи |
40 |
|
Сердце быка |
8—50 |
Пшеничная мука |
9—25 |
|
Яйцо (желток) |
30 |
Цветная капуста |
17 |
|
Сардины (консервы) |
24 |
Шампиньоны |
16 |
|
Ветчина |
10 |
Горошек зеленый |
6 |
|
Камбала |
8 |
Бананы |
4,4 |
|
Мясо курицы |
6 |
Дыня |
3,6 |
|
» говядины |
5 |
Лук свежий |
3,5 |
|
Молоко коровье |
5 |
Капуста белокочанная |
2.5 |
|
Сыр |
4 |
Морковь |
2,5 |
|
Сельдь |
4 |
Апельсины |
1,9 |
|
Телятина |
1,5—2 |
Яблоки |
0,9 |
|
Соя, бобы |
60 |
Картофель |
0,5—1,0 |
Наиболее богаты витаминами свиная и говяжья печень, почки, сердце быка, яичный желток, а из продуктов растительного происхождения—бобы, рисовые отруби, пшеничная мука и цветная капуста. В животных тканях и дрожжах биотин находится преимущественно в связанном с белками виде, в овощах и фруктах—в свободном состоянии.
4. Биосинтез биотина.
Биосинтез биотина осуществляют все зеленые растения, некоторые бактерии и грибы. Изучение путей биосинтеза биотина началось после выяснения строения его молекулы. Химическое расщепление биотина проходит через образование дестиобиотина, диаминопеларгоновой кислоты и, наконец, пимелиновой кислоты. Вполне естественно было предположить, что биосинтез биотина может проходить путем постепенного усложнения молекулы пимелиновой кислоты. В пользу этого говорил тот факт, что пимелиновая кислота способна заменять биотин у некоторых микроорганизмов как фактор роста. Она стимулирует синтез биотина: меченая пимелиновая кислота обнаруживается в углеродном скелете биотина.
Изучение структурной формулы биотина привело к предположению, что атомы I,1', 4 и 5 происходят из декарбоксилированной молекулы цистеина (см. формулу), атомы 2 и 3 происходят из карбамилфосфата и связаны, таким образом, с имеющимся в клетках «пулом» СО2 в то время как остальные семь атомов (2, 3, 6, 7, 8, 9 и 10) происходят из углеродного скелета пимелиновой кислоты. Высказанное предположение подтверждено экспериментально при изучении биосинтеза
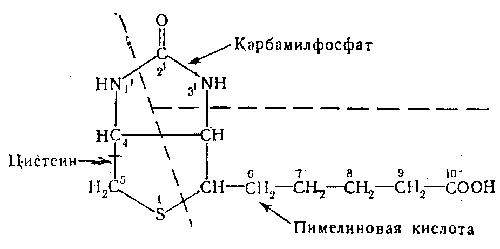
Биотина в культурах Achromobacter, выращенных на синтетических средах, к которым добавляли либо 3-С14-цистеин, либо МаНС14Оз. Синтезированный бактериями радиоактивный биотин расщепляли и таким образом изучали распределение в нем радиоактивного углерода. Основываясь на полученных результатах, Lezius и соавторы в 1963 г. предложили схему синтеза биотина.
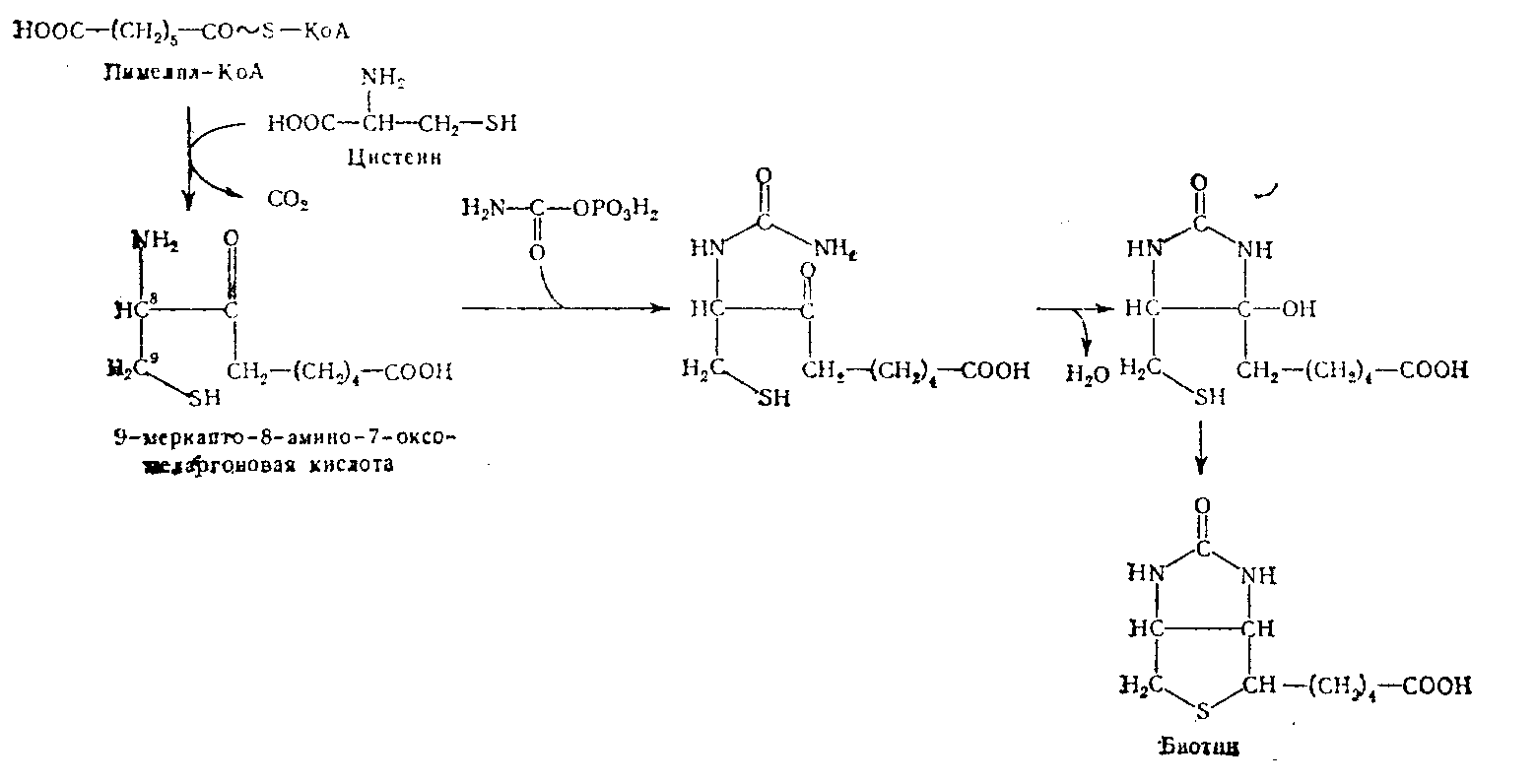
Согласно этой схеме, началом синтеза является конденсация пимелил-КоА и цистеина. Затем происходит декарбоксилирование, что приводит к образованию 9-меркапто-8-амино-7-оксопеларгоновой кислоты. Взаимодействие аминогруппы этой кислоты с карбамилфосфатом вызывает образование уреидного производного, которое после отщепления воды может циклизоваться, давая биотин с характерным для него двойным циклом.
По способности синтезировать биотин и дестиобиотин все исследованные организмы делятся на 4-е группы:
Способные синтезировать большое количество биотина и дестиобиотина из глюкозы в отсутствие пимелиновой кислоты.
Стимулирующие при помощи пимелиновой кислоты и дестибиотина биосинтез биотина.
Активно осуществляющие превращение дестибиотина в биотин.
Образующие дестиобиотин из пимелиновой кислоты, но не способные превращать его в биотин.
Изучено более 600 штаммов бактерий, использующих углеводороды для синтеза биотина, из которых 35, синтезируют витамин в больших количествах (>100 мкг/мг). Наибольшее количество биотина образует Pseudomonas sp. штамм 5-2 при выращивании на керосине. Специфическим активатором накопления биотина является аденин. Экзогенные пимелиновая и азелаиновая кислоты увеличивают образование блотина. из керосина. Лучшими источниками углерода оказались н-алканы с углеродной цепью из 15—20 атомов, в частности н-ундекан. Промежуточными продуктами в синтезе биотина из ундекана являются пимелиновая и азелаиновая кислоты (Toshimichi e. a., 1966).
Исследование биосинтеза биотина в растениях (В. Филиппов, 1962 г.) показало, то каждый орган растения и каждая его клетка синтезирует витамин в эмбриональной фазе своего развития. В дальнейшем синтез замедляется и, по-видимому, прекращается, но содержание его различных тканях долгое время остается постоянном.
5. Обмен биотина в организме
Об обмене биотина известно немного. Биотин, поступивший с пищей в связанном состоянии, отщепляется от белка под действием протеолитических ферментов, переходит в водорастворимую форму и всасывается в кровь в тонком кишечнике. В кишечнике происходит также всасывание биотина, синтезированного бактериями желудочно-кишечного тракта. Всосавшийся в кровь биотин связывается с альбумином сыворотки разносится по всему организму. Наибольшее количество биотина накапливается в печени, почках и надпочечниках, причем у мужчин оно несколько больше, чем у женщин.
Содержание биотина в тканях человека (Р. Д. Вильяме, 1950)
|
Органы и ткани |
Биотин в мкг/г |
|
|
у женщин |
у мужчин |
|
|
Кожа |
— |
0 01 |
|
Мозг |
0,03 |
0,08 |
|
Легкие |
0,02 |
0,01 |
|
Сердце |
0,17 |
0,19 |
|
Мышцы |
0,02 |
0,04 |
|
Желудок |
0,19 |
0,11 |
|
Ободочная кишка |
0,08 |
0,09 |
|
Печень |
0,62 |
0,77 |
|
Молочная железа |
0,04 |
— |
|
Селезенка |
0,04 |
0.06 |
|
Почки |
0 58 |
0,67 |
|
Надпочечники |
0,35 |
0,23 |
|
Семенники |
— |
0,05 |
|
Яичники |
0,03 |
—— |
Что касается содержания биотина в крови человека, то по этому вопросу имеется ограниченная и порой противоречивая информация. Bhagavan и Coursin в 1967 г. определили содержание биотина микробиологическим методом в крови 30 здоровых лошадей и 25 взрослых людей и показали, что в среднем в крови взрослых людей содержится 25,7 ммкг% биотина (12—42,6 ммкг%), а в крови детей несколько больше—32,3 ммкг% (14,7—55,5 ммкг%). По данным Baugh (1968), средний уровень биотина в цельной крови составляет 147 ммкг% (82— 270 ммкг%). Какой-либо разницы, в содержании биотина в .крови в зависимости от пола и возраста не отмечено. Содержание биотина в молоке женщины резко изменяется в период кормления. В первый день после родов содержание биотина b молоке невелико и только на 10-й день повышается до 0,33 мкг на 100 мл.
Биотин почти не подвергается Обмену в организме человека и выводится в неизмененном виде в основном с мочой. У здоровых людей выведение биотина с мочой составляет 11—183 мкг в сутки, у новорожденных детей достигает максимума (4 мкг на 100 мл)_ на 2-й день жизни и снижается до нуля к 7-му дню. Содержание биотина в кале колеблется от 322 до 393 mкг в сутки. В норме выделение биотина с мочой и калом повышает поступление его с пище 3-6 раз. что свидетельствует о удовлетворении потребностей человека в биотине на счет бактериального синтеза в кишечнике. Через 6 часов после введения человеку массированной дозы биотина большая часть его выводится с мочой. Содержание биотина в кале при этих же условиях изменяется в меньшей степени.
Небольшая часть карбоксильной группы боковой цепи биотина окисляется до СО2 специфической оксидазой, которая обнаружена в печени и почках морской свинки и крысы.
Исследование распределения меченого биотина в тканях цыплят и крыс показало, что уже через 4 часа после выведения физиологической дозы меченного С1' по карбоксильной группе биотина около 16% метки включалось в печень, а 30% выводилось с калом и мочой в неизмененном виде (Dakshinamurty, Mistry, 1963). В сердце, селезенке и легких радиоактивности не обнаружено. Менее 4% введенной дозы выводилось в виде выдыхаемого C14O2, что указывало на незначительное прямое окисление карбоксильной группы биотина. О распределении меченого биотина в различных клеточных фракциям можно судить по табл.
Содержание биотина в клеточных фракциях печени нормальных крыс (Dakshinamurti, Misfry, 1963)
Фракция печени |
Нормальные животные |
Авитаминозные животные |
|
|
Общий биотин в % |
Связанный биотин в % к общему |
Общий биотин (в %) |
|
|
Гомогенат |
100 |
92 |
100 |
|
Ядра |
37 |
99 |
75 |
|
Митохондрии |
9 |
89 |
13 |
|
Микросомы |
2 |
23 |
о |
|
Надосадочная жидкость |
47 |
91 |
о |
Из таблицы видно, что 40—50% радиоактивности обнаружено в надосадочной фракции, полученной после центрифугирования гомогената печени крыс. В микросомах содержится незначительное количество витамина. Большая часть биотина в различных клеточных фракциях, за исключением микросом, присутствует в связанной с белком форме. Имеются и противоречивые данные о том, что большая часть биотина (более 60%) содержится в митохондриях печени животных и около 11%— в микросомах.
В настоящее время недостаточно исследована динамика содержания биотина в тканях в онтогенезе животных. По-видимому, яйцо и зародыш в начальной стадии развития наиболее богаты биотином. Развитие зародыша сопровождается снижением содержания биотина в тканях. Исключение составляют печень и почки, в которых содержание биотина значительно повышается в первые дни постэмбрионального развития.
6. Участие биотина в обмене веществ и механизм действия
К 1958—1959 гг. накопились данные, которые указывали на участие биотина в реакциях карбоксилирования. Установлено, что при биотиновой недостаточности нарушаются следующие функции печени животных:
синтез цитруллина из орнитина, МН3 и С02, включение CО2 в пурины, карбоксилирование пропионовой кислоты, приводящее к образованию янтарной кислоты, включение С02 в ацетоуксусную кислоту. Однако механизм действия биотина в этих реакциях оставался невыясненным. Данные опытов с 2-C14-биoтинoм исключали возможность того, что С-атом уреидной группировки биотина переносится в качестве остатка угольной кислоты. Одним из обстоятельств, из-за которых подвергалась сомнению функция этого витамина как кофермента карбоксилирования, было (описанное в разное время) участие биотина в реакциях, в которых не происходило ни включения, ни отщепления С02. Так, было обнаружено влияние биотина на дезаминирование аспарагиновой кислоты, серина и треонина и участие его в синтезе жирных кислот. Первые четкие доказательства коферментной функции биотина в реакции карбоксилирования появились в работах, посвященных именно синтезу жирных кислот. В этих работах отмечалось, что биотин является коферментом ацетил-КоА-карбоксилазы, фермента, осуществляющего карбоксилирование ацетил-КоА с образованием малонил-КоА—первую стадию синтеза жирных кислот (Wakil, 1958). К этому времени были получены доказательства существования еще одного биотинфермента, а именно (З-метил-кротонил-КоА-карбоксилазы (Lynen, Knappe, 1959). Все известные в настоящее время биотиновые ферменты катализируют два типа реакций:
Реакции карбоксилирования или фиксации С02, сопряженные с расщеплением АТФ и протекающие согласно уравнению:
АТФ +
НСОз + RH![]() R—СОО-
+ АДФ + Фнеорг.
R—СОО-
+ АДФ + Фнеорг.
|
Реакции |
Источник фермента |
|
Ацетил-КоА
+ С02+
АТФ
Я-Метилкротонил-КоА
+ С02
+ АТФ
Пропионил-КоА
+ С02
+ АТФ
Бутирил-КоА+С02
+ АТФ
Пируват
+ С02+
АТФ |
Печень голубя Микробактерии Сердце и печень свиньи Мышцы и печень голубя Печень голубя |
II. Реакции транскарбоксилирования, протекающие без распада АТФ, при которых карбоксилирование одного субстрата осуществляется при одновременно протекающем декарбоксилировании другого соединения:
R1—COO-
+ R2H![]() R1H
+ R2—
COO-
R1H
+ R2—
COO-
Поскольку все приведенные реакции являются обратимыми, возможен обратимый биосинтез АТФ. Во всех этих случаях имеет место включение С02 в реактивное α-подожение ацил-КоА или винилгомоло-гичное ему положение (при карбо^силировании β-метилкротонил-КоА).
К началу 60-х год5В были выделены и изучены карбоксилазы, осуществляющие указанные превращения —В 1960 г. установлено участие биотина в реакции транскарбоксилирования при исследовании синтеза пропионовои кислоты
СНз—СН—СО~S—КоА
+ СНз—СО—СООН
![]()
![]()
СООН
![]() СНз-СНа—СО~S-КоА
+ НООС-СН2—СО—СООН
СНз-СНа—СО~S-КоА
+ НООС-СН2—СО—СООН
Биотиновые ферменты представляют собой олигомеры с большим молекулярным весом (порядка 700000) и, как правило, содержат 4 моля связанного биотина на 1 моль фермента, поэтому кажется вероятным, что они состоят из 4 субъединиц с молекулярным весом 175000, каждая из которых содержит одну молекулу биотина.
В работах Lynen (1964) расшифрован механизм участия биотина в реакциях карбексилирования. Установлено, что реакции карбоксилирования являются двухстадийными. Первая стадия сводится к образованию «активной С02» в форме С02~биотинфермента:
АТФ + Н
С0-2+
биотинфермент
![]() АДФ
+ Фнеорг. + С02~биотинфермент.
АДФ
+ Фнеорг. + С02~биотинфермент.
Вторая стадия заключается в переносе «активной С02» на акцептор:
С02~биотинфермент
+ R2H
![]() биотинфермент
+ R2—
С00-
биотинфермент
+ R2—
С00-
Аналогичный двух стадийный механизм предложен и для реакций транскарбоксилирования:
R1
—С00-
+ биотинфермент
![]() С02
~биотинфермент
R2H;
С02
~биотинфермент
R2H;
С02~биотинфермент
+ R2H![]() R2—
С00-
+
биотинфермент.
R2—
С00-
+
биотинфермент.
После установления существования «активной С02» в виде С02~биотинфермента установлен характер связи между С02 и биотином. Этому способствовало открытие того факта, что β-метилкротонил-КоА-карбоксилаза способна карбоксилировать свободный биотин, переводя его в карбоксибиотин. В дальнейшем меченый карбоксибиотип был выделен в опытах с С14-бикарбонатом и идентифицирован как Г-М-карбоксибиотин. Его структура была подтверждена химическим синтезом. К атому времени уже было известно, что в биотиновых ферментах карбоксильная группа биотина соединена с ε-NH2-группой лизина ферментного белка ковалентной связью. На основании этих данных предложена структура С02~биотинфермента.
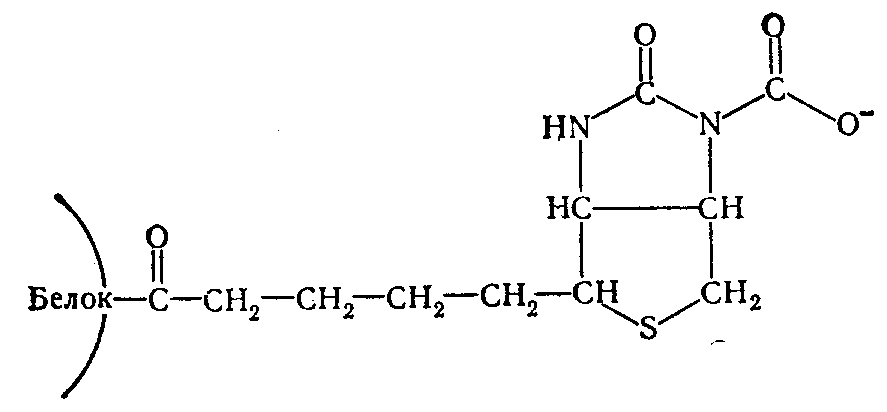
Эта структура получила ряд экспериментальных подтверждений и в настоящее время является общепринятой для всех биотиновых ферментов. Реакционная способность углекислоты, связанной с биотином, находит выражение в энергетических взаимоотношениях. Величина свободной энергии распада С02~биотинфермента равна 4,74 ккал/моль, что дает основание причислить С02~биотинфермента к «богатым энергией» соединениям.
Исключительно большой интерес представляет совершенно неизученная проблема регуляции активности биотинсодержащих ферментов и организме. В этой связи особенно важны исследования по биосинтезу молекулы биотина и образованию холоферментов из биотина и соответствующего ферментного белка. Данные по первому вопросу изложены в разделе «Биосинтез». Что касается образования холофермента, то можно считать установленным, что во всех биотиновых ферментах био-тнн связан с ε-аминогруппой лизина. Этот способ связи экспериментально доказан почти для всех карбоксилаз и метилмалонил-КоА-оксалоаце-таттранскарбоксилазы. Недостаточные по биотину клетки Propionibacterium shcemanii содержат апофермент и специфическую синтетазу, которая катализирует при использовании АТФ соединение биотина с апоферментом, приводящее к образованию активного холофермента траискарбокснлазы. Необходимыми кофакторами этой реакции являются АТФ и Mg2+. При использовании очищенных ферментов удалось доказать, что образование холотранскарбоксилазы происходит в два этапа, причем промежуточным соединением является биотиниладенилат (R-CO-5'-AMФ):
Mg2+
I. АТФ
+ R— С02Н
+ синтетаза![]() R-СО-5’-
АМФ
-
синтетаза
+ пирофосфат
R-СО-5’-
АМФ
-
синтетаза
+ пирофосфат
(биотин
I![]() I.
R-СО-5’-
АМФ
-
синтетаза
+ Н2М-фермент
R-CO-NH-фермент +
I.
R-СО-5’-
АМФ
-
синтетаза
+ Н2М-фермент
R-CO-NH-фермент +
+5’-AMФ+cинтeтaзa.
Синтетический биотиниладенилат обладает способностью заменить смесь АТФ, MgCl и биотина при синтезе холофермента (Lynen, 1964). Позже было установлено, что образование других холоферментов протекает аналогичным образом. Все известные ферментативные реакции, для которых установлено участие биотина в качестве кофермента, являются процессами переноса углекислоты. По-видимому, в обратимом присоединении и отдаче СО; и состоит исключительная функция этого витамина в обмене веществ. Однако при биотиновой недостаточности нарушаются очень многие реакции обмена в интактном организме. Так, - биотин вовлечен в биосинтез белков, дезаминирование аспартата, серина и треонина у бактерий, обмен триптофана, жиров и углеводов, синтез пуринов, образование мочевины у животных и др. Природа участия биотина во многих из этих реакций остается неясной. Все перечисленные процессы имеют одну общую черту: при изучении in vitro они не тормозятся авидином. На основании этих данных считается, что биотин оказывает.непрямое действие на указанные превращения, которые катализируются ферментами, не содержащими этого витамина.
Ввиду чрезвычайной важности нeкоторых из этих реакций для жизне-деятельности организма необходимо рассмотреть их. Рядом авторов отмечено, что при недостаточности биотина в рационе крыс снижается включение в белок меченых аминокислот. Так, включение (С14-метионина, С14-лейцина и С14-лизина в тканевые белки снижается на 20—40% причем недостаточность биотина влияет на стадию образования амино-ацил-транспортной РНК. Препараты тРНК из печени нормальных крыс включают значительно больше меченых аминокислот, чем препараты печени авитаминозных животных (Dakshinainurti, Misty, 1964). Еще ранее было установлено, что у авитаминозных животных нарушается синтез амилазы в поджелудочной железе и сывороточного альбумина в печени, причем однократное введение 100 мкг биотина восстанавливает способность тканей к синтезу указанных белков. Добавление in vitro α-кетоглутарата и фумарата также восстанавливает образование амилазы и сывороточного альбумина (А. А. Познанская, 1957).
Эти данные показали, что биотин не принимает прямого участия в синтезе белка de novo, а его влияние на этот процесс, по-видимому, определяется вовлечением биотина в синтез субстратов трикарбонового цикла. Такое предположение нашло подтверждение в опытах на цыплятах: скармливание сукцината авитаминозным птицам восстанавливало до нормы включение аминокислот в тканевые белки и РНК. Так как образование С4-дикарбоновых кислот в организме животных протекает через фиксацию СО2, осуществляемую биотиновыми ферментами, то становятся ясными причины нарушения синтеза белка при биотиновой недостаточности. В организме авитаминозных цыплят значительно снижены скорость окисления глюкозы до СО2 и включение ее в гликоген печени. Имеются указания на снижение глюкокиназной активности при недостаточности биотина, хотя витамин не был обнаружен в препаратах кристаллического фермента. Возможно, что участие биотина в обмене. углеводов является непрямым. В результате нарушения утилизации глюкозы в организме животных при исключении биотина из корма нарушается превращение D-глюкозы в L-аскорбиновую кислоту.
При недостаточности биотина содержание липидов в печени животных снижается на 30°/о, что обусловлено снижением синтеза жирных кислот. Отсутствие биотина в корме цыплят приводит к повышению содержания триглицеридов, пальмитиновой и пальмитолеиновой кислот в печени, а также соотношения жирных кислот СО16, СО18 соотношение между насыщенными жирными кислотами при этом снижается. Включение введенных СО14-стеариновой и СО14-пальмитиновой кислот в фосфолипиды значительно повышается при недостаточности биотина, а включение их в триглицериды—снижается. При этом общее количество жирных кислот по сравнению с контролем понижено. В печени авитаминозных крыс снижено включение СО14-ацетата в липиды и наблюдается большее содержание ненасыщенных жирных кислот СО16:1 и СО18:2, тогда как уровень стеариновой кислоты снижен. Исключение биотина из корма крыс в течение 60 дней приводит к значительному снижению уровня цитидиловых, адениловых и гуаниловых нуклеотидов в печени. Содержание уридиловых нуклеотидов при этом почти не изменяется, а инозиловых — немного повышается. В то же время отсутствие биотина в рационе не влияет на содержание РНК и ДНК, а также на включение в них Р32, введенного внутрибрюшинно в виде Na2HP32O4.
Наконец, в гомогенатах печени авитаминозных крыс резко снижено образование цитруллина, которое полностью восстанавливается через 24 часа после введения животным биотина. Однако биотин не обнаружен в препаратах ферментов, участвующих в синтезе мочевины (карбамилфосфатсинтетаза, орнитинтранскарбамилаза и др.) и, по-видимому, в данном случае оказывает непрямое действие на эту реакцию.
6.1 Взаимодействие с другими витаминами. Установлена связь биотина с другими витаминами, в частности с фолиевой кислотой, витамином B12 - аскорбиновой кислотой, тиамином и пантотеновой кислотой. 0собенно тесные взаимоотношения существуют между биотином и фолиевой кислотой. Сначала было показано, что при недостатке биотина в печени крыс значительно снижено общее содержание веществ, обладающих активностью фолиевой кислоты. и что биотин стимулирует биосинтез этого витамина с флорой. Позднее было установлено, что у биотинавитаминозных крыс значительно снижено содержание коферментны.х форм фолиевой кислоты, а именно N5 и N10-формилтетрагидро-фолатов, тетрагидрофолата, N5_ и N10_ формилтетрагидроптероилглутаминовых кислот. Биотин стимулирует синтез метионина из серина и гомоцистеина и процессы метилирования вообще, способствуя накоплению коферментных форм фолиевой кислоты. Таким образом, при недостаточности биотина нарушена утилизация организмом фолиевой кислоты и превращение ее в активные коферментные формы. По-видимому, биотин принимает непосредственное участие в ферментативных процессах превращения фолиевой кислоты в ее коферментные производные (Marchetti e. а., 1966). Биотин благоприятно влияет на общее состояние организма и сохранение аскорбиновой кислоты в тканях цинготных морских свинок. В свою очередь аскорбиновая кислота замедляет, хотя и не предотвращает развитие авитаминоза биотина у крыс. При недостаточности биотина снижается содержание тиамина в печени, селезенке,. почках и мозге животных. У крыс, содержавшихся на рационе, лишенном биотина, содержание витамина B12 было выше, чем у контрольных животных, получавших биотин. Эти два витамина тесно связаны между собой в обмене пропионовой кислоты у микроорганизмов и животных. Существует тесная связь между биосинтезом биотина и пантотеновой кислоты у микроорганизмов и зеленых растений (В. В. Филиппов, 1962). Биотин облегчает симптомы пантотеновой недостаточности и, наоборот, пантотеновая кислота смягчает проявление авитаминоза биотина.
7. Потребность организма в биотине.
Биотин необходим для человека, животных, растений и большого числа микроорганизмов. Он является фактором роста для многих штаммов, а также многих грибов и бактерий. Однако некоторые дрожжи, грибы и бактерии способны его синтезировать. Потребность в биотине у птиц и животных покрывается за счет синтеза его бактериями желудочно-кишечного тракта. У коров, овец и лошадей, содержащихся на обычном рационе, практически исключена недостаточность биотина. У свиней и птиц недостаточность биотина может создаваться при использовании кормов, бедных витаминами.
Потребность в биотине у человека покрывается за счет синтеза его микрофлорой кишечника, поэтому ее трудно оценить. С известной долей приближения можно считать, что (минимальной ежедневной дозой биотина для животных и человека являются следующие величины (Gyorgy, 1954): для человека— 150—200 мкг, обезьян—20 мкг, крыс— 0,5—3 мкг, цыплят — 0,65—1 мкг, свинец — 100 мкг.
В период беременности и лактации Потребность в биотине у женщин повышается до 250— 300 мкг в день. По другим данным, потребность в биотине значительно ниже и составляет для взрослого человека 30— 40 мкг в сутки.
7.1 Проявление недостаточности биотина
Наиболее подробно недостаточность биотина изучена в опытах на крысах и цыплятах при скармливании рационов с большим содержанием сырого яичного белка. Биотиновый авитаминоз у животных характеризуется прекращением роста и падением веса тела (до 40%), покраснением и шелушением кожи, выпадением шерсти или перьев, образованием красного отечного ободка вокруг глаз в виде «очков», атактической походкой, отеком лапок и типичной позой животного с согбенной (кенгу-руподобной) спиной. Дерматит, который развивается у животных при недостаточности биотина, может быть охарактеризован как себорея десквамационного типа, сходная с той, которая наблюдается у детей. У крыс авитаминоз биотина развивается через 4—5 недель скармливания опытного рациона, а у цыплят первые признаки авитаминоза появляются через 3 недели.
Помимо внешних признаков, биотиновый авитаминоз вызывает глубокие морфологические изменения в тканях и органах, а также нарушения в обмене веществ. Известны изменения в зобной железе, коже и мышцах крыс. Характерны обильный гиперкератоз, акантез и отеки. Разрушенные волосяные стволы перемешаны с гиперкератозными пластинками. Установлено расширение волосяных сумок, отверстия которых закупорены гиперкератозным материалом. В последней фазе развития авитаминоза наблюдается атрофия жира в гиперкератозных пластинках. Недостаток биотина в рационе крыс приводит к уменьшению его содержания в тканях. В печени и мышцах количество витамина снижается в 5 раз, а в мозговой ткани—на 15%. В крови авитаминозных крыс накапливается пировиноградная кислота, развивается ацидоз и снижается концентрация сахара. При этом глюкозурия не наблюдается, но уменьшается содержание редуцирующих Сахаров в печени при нормальном содержании их в мышцах; у животных развивается креа-тинурия.
Человек полностью удовлетворяет свою потребность в биотине за счет синтеза его микрофлорой кишечника, поэтому гиповитаминоз можно получить только в эксперименте. Экспериментальную недостаточность биотина у человека наблюдали Sydenstricker и соавторы (1942) путем включения в диету ежедневно 200 г сырого яичного белка. Через 3 недели появилось шелушение кожи без зуда. На 7—8-й день развилась пепельная бледность кожи и началась атрофия вкусовых сосочков языка. Позднее появились мышечные боли, повышенная чувствительность, болезненные ощущения, вялость, сонливость, тошнота и потеря аппетита. В крови уменьшилось содержание эритроцитов и холестерина. Выделение биотина с мочой снизилось в 7—8 раз против нормы (с 29— 52 до 3,5—7,3 мкг в сутки). Введение 150 мкг биотина уже на 3—4-й день устраняло депрессию, мышечные боли и восстанавливало аппетит.
Таким образом, недостаточность биотина у человека в первую очередь вызывает поражения кожи. Более тяжелые проявления требуют, по-видимому, большей длительности авитаминоза.
8. Профилактическое и лечебное применение биотина.
В настоящее время применение биотина в клинике с лечебной и профилактической целью изучено недостаточно. Большое значение биотина для нормального состояния кожных покровов привело к попыткам лечения биотином ряда кожных заболеваний. Биотин показан при себорейном дерматите у грудных детей, связанном, возможно, с явлениями недостаточности этого витамина. Заболевание излечивается при ежедневном введении 5—10 мкг биотина в течение 4 недель. Отмечен успех при длительном применении биотином в тех случаях дескваматозной эритродермии, когда недостаточность биотина была в числе причин заболевания. Лечебная доза биотина составляет 150—300 мкг в сутки; вводится он-парентерально.
Г. И. Бежанов в 1966 г. сообщил о применении биотина в комплексной терапии псориаза. Наблюдения, проведенные за большой группой больных, показали, что биотин проявлял противозудный эффект, а сочетание биотина с фумаратом, витаминами группы В и бальнеотерапией потенцированный и более быстрый клинический эффект. В процессе лечения у значительного большинства больных рассасывался инфильтрат, уменьшалось или полностью прекращалось шелушение. Автор рекомендует применять биотин в комплексном лечении псориаза.
В последние годы появился ряд сообщений о целесообразности применения фармакологических доз биотина п комплексной терапии атеросклероза и гипертонической болезни.. Эти данные представляют особый интерес в связи с участием биотина в синтезе холестерина.
О. К. Докусова и А. Н. Климов в 1967 г. сообщили о предотвращении биотином экспериментального атеросклероза у кроликов при скармливании больших доз холестерина. Введение биотина в количестве 400 мкг в день предупреждало развитие атеросклероза. Содержание холестерина, β -липопротеидов и фосфолипидов в стенке аорты животных, получавших холестерин и биотин, не отличалось от содержания этих компонентов в аорте здоровых кроликов, тогда как в аорте животных, получавших холестерин без биотина, содержание холестерина и β-липопро-теидов было резко повышено. Авторы считают, что снижение содержания холестерина вызвано окислением его в печени. В настоящее время известно, что пропионат является основным трехуглеродным фрагментом, отщепляющимся от боковой цепи холестерина на первых этапах окисления холестерина в желчные кислоты. Можно предположить, что стимуляция окисления холестерина биотином связана с активацией окисления пропионовой кислоты (через образование янтарной кислоты путем кар-боксилирования пропионовой кислоты), поскольку пропионил-КоА-кар-боксилаза является биотинсодержащим ферментом.
В. Д. Устиловский и др. (1967) сообщил, что после 7-дневного введения внутрь фармакологических доз биотина у больных атеросклерозом, отмечалось статистически достоверное снижение содержания общего, холестерина и β -липопротеидов в крови, тогда как у здоровых людей биотин не влиял на изучаемые показатели. У больных атеросклерозом и гипертонической болезнью применение фармакологических доз биотина приводит к значительному снижению выведения с мочой тиамина и аскорбиновой кислоты (М. А. Лис, 1967; Д. П. Калкун, 1967).
Таким образом, биотин способствует усвоению этих витаминов в организме больных. Учитывая благоприятное влияние биотина на ряд показателей липидного обмена у больных атеросклерозом и гипертонической болезнью, а также повышение усвоения тиамина и аскорбиновой кислоты, авторы рекомендуют применять биотин при этих заболеваниях в дозе 1 мг в день внутрь в комплексной терапии,
С возрастом в крови людей снижается содержание пантотеновой кислоты, витамина В6 и биотина (В. И. Титов, 1966). У больных гипертонией и атеросклерозом в возрасте 80—88 лет содержание биотина было в 2,4 раза меньше, чем у здоровых молодых людей. Установлено снижение содержания биотина в крови больных, жаловавшихся на слабость, недомогание и легкую утомляемость. Автор рекомендует применять пантотеновую кислоту, витамин В6 и биотин при преждевременной старости для устранения явлений гиповитаминоза и связанного с ним нарушением обмена веществ. Некоторыми исследователями обнаружено высокое содержание биотина в тканях ряда раковых опухолей, что привело к попыткам воздействовать на развитие рака путем вызывания биотиновой недостаточности. Однако эти попытки не дали положительных результатов.
Таким образом, сейчас накапливаются данные, позволяющие рекомендовать применение биотина при некоторых болезнях кожи, нарушениях жирового обмена и сердечно-сосудистых заболеваниях. Однако для более широкого использования биотина в клинике необходимо дальнейшее изучение как его физиологического действия, так и показаний к лечебному применению.
Тиамин
Исторические сведения
Первые упоминания о заболевании (какке, бери-бери), известном сейчас как проявление недостаточности тиамина, встречаются в древних медицинских трактатах, дошедших до нас из Китая, Индии, Японии (Bicknell, Prescott, 1953; Inouye, Katsura, 1965). К концу прошлого столетия клинически уже различали несколько форм этой патологии, но только Takaki (1887) связал заболевание с какой-то, как он тогда полагал, недостаточностью азотсодержащих веществ в пищевом рационе. Более определенные представления были у голландского врача С. Eijkman (1893—1896), обнаружившего в рисовых отрубях и в некоторых бобовых растениях неизвестные тогда факторы, предупреждавшие развитие или излечивавшие бери-бери. Очисткой этих веществ занимались затем Funk (1924), впервые предложивший сам термин «витамин», и ряд других исследователей (Wuest, 1962). Извлеченное из естественных источников активное вещество только в 1932 г. было охарактеризовано общей эмпирической формулой, а затем (1936) успешно синтезировано. Еще в 1932 г. высказывалось предположение о роли витамина в одном из конкретных процессов обмена веществ—декарбоксилировании пировиноградной кислоты, но лишь в 1937 г. (Lohman, Schuster) стала известна коферментная форма витамина—тиаминдифос-фат (ТДФ). Коферментные функции ТДФ в системе декарбоксилирования ct-кето-кислот долгое время представлялись почти единственными биохимическими механизмами реализации биологической активности витамина, однако уже в 1953 г. круг ферментов, зависящих от присутствия ТДФ, был расширен за счет транскетолазы, а совсем недавно и специфической декарбоксилазы γ-окси-α-кетоглютаровой кислоты. Нет оснований думать, что перечисленным исчерпывается перспектива дальнейшего изучения витамина, так как эксперименты на животных, данные, получаемые в клинике при лечебном применении витамина, анализ фактов, иллюстрирующих известную нейро- и кардиотропность тиамина, с несомненностью указывает на наличие еще каких-то специфических связей витамина с другими биохимическими и физиологическими механизмами (В. Б. Спиричев, 1966; Ю. М. Островский, 1971).
2. Химические и физические свойства витамина В1
Тиамин (В. М. Березовский, 1959; В. А. Девятнин, 1964), или 4-метил-5-β-оксиэтил-N- (2-метил-4-амино-5-метилпиримидил) -тиазолий, получается синтетически обычно в виде хлористо-или бромистоводородной соли:
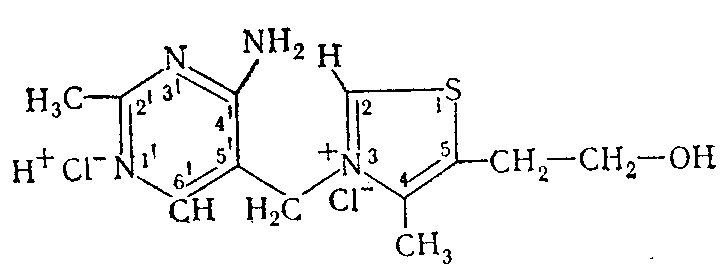
-Рйаминхлорид (М-337,27) кристаллизуется с Ѕ Н2О в бесцветных моноклинических иглах, плавится при 233—234° (с разложением). В нейтральной среде его спектр поглощения имеет два максимума — 235 и 267 нм, а при рН 6,5 Один — 245—247 нм. Витамин хорошо растворяется в вода и уксусной кислоте, несколько хуже в этиловом и метиловом спиртах и нерастворим в хлороформе, эфире, бензоле, ацетоне. Из водных растворов тиамин может быть осажден фосфорно-вольфра-мовой или пикриновой кислотой. В щелочной среде тиамин подвергается многочисленным превращениям (Metzler, 1960), которые, в зависимости от природы добавленного окислителя, могут завершаться образованием тиаминдисульфида (X) или тиохрома (IX).
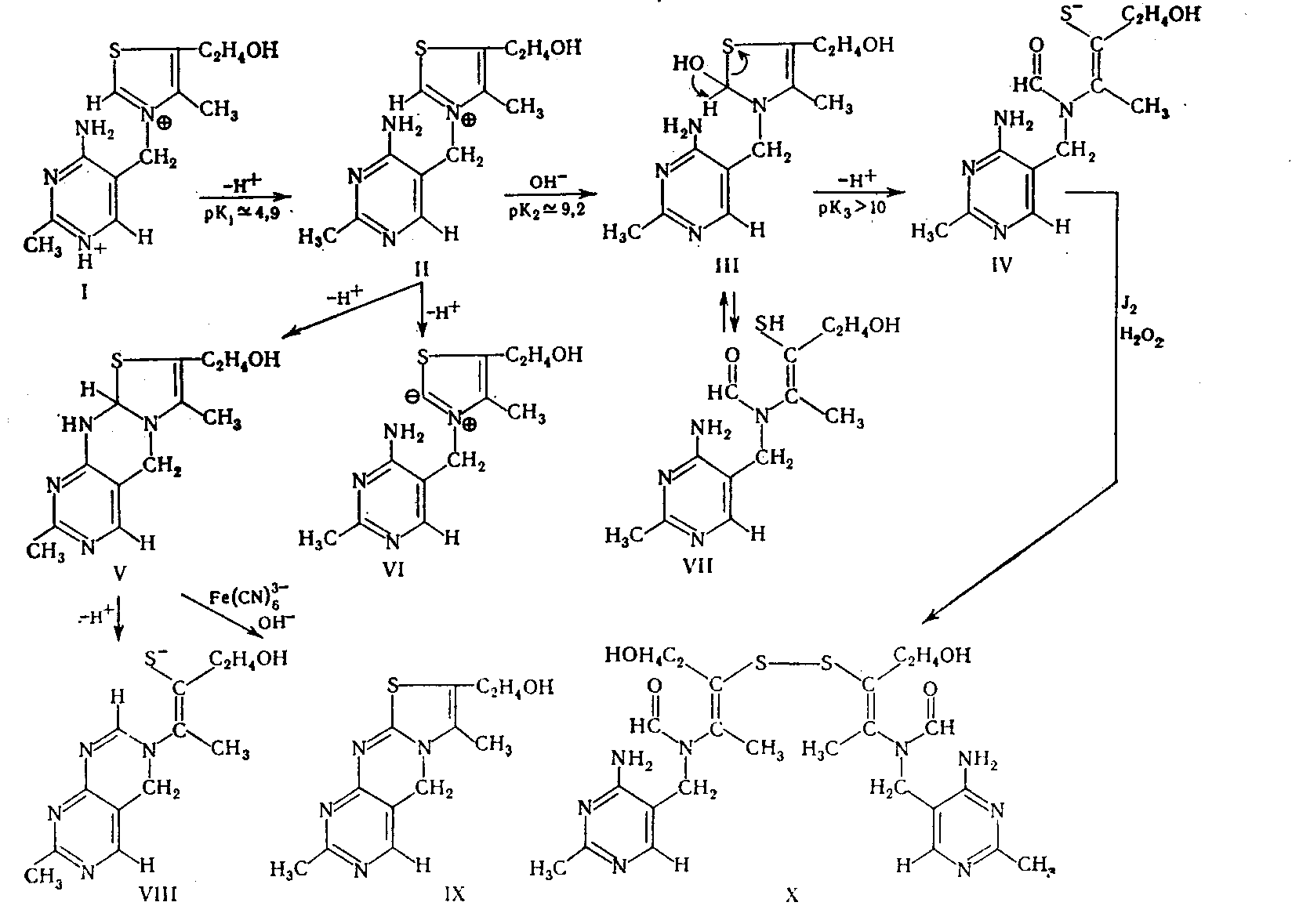
В кислой среде витамин разлагается только при длительном нагревании, образуя 5-гидрокси-метилпиримидин, муравьиную кислоту, 5-аминометилпиримидин, тиазоловый компонент витамина и З-ацетил-3-меркапто-1-пропанол. Среди продуктов распада витамина в щелочной среде идентифицированы тиотиамин, сероводород, пиримидодиазепин и др. Получены также сульфат и мононитрат витамина. Известны соли тиамина с нафталенсульфоновой, арилсульфоновой, цетилсерной и эфиры с уксусной, пропионовой, масляной, бензойной и другими кислотами.
Особое значение имеют эфиры тиамина с фосфорной кислотой, в частности ТДФ, являющийся коферментной формой витамина. Получены (Fragner, 1965; Schellenberger, 1967) гомологи тиамина путем различных замещений у второго (этил-, бутил-, оксиметил-, оксиэтил-, фенил-, оксифенил-, бензил-, тиоалкил-), четвертого (окситиамин) и шестого (метил-, этил) атомов углерода пиримидина метилированием аминогруппьь, замещением тиазоловогоинхла на пиридиновой (пиритиамин), имидозоловый или оксазфювый, модификациями заместителей у пятого углерода тиазола (метил-, оксиметил-, этил-, хлорэтил-, оксипропил- и др.). Отдельную большую группу соединений витамина составляют S-алкильные и дисульфидные производные (Matsukawa e. а., 1970). Среди последних наибольшее распространение как витаминный препарат получил тиаминпропилдисульфид (ТПДС).
3. Распространение витамина В1, в природе.
Тиамин распространен повсеместно и обнаруживается у разных представителей живой природы (Р. В. Чаговец и др., 1968). Как правило, количество его в растениях и микроорганизмах достигает величин значительно более высоких, чем у животных. Кроме того, в первом случае витамин представлен преимущественно свободной, а во втором — фосфорилированной формой. Содержание тиамина в основных продуктах питания колеблется в довольно широких пределах в зависимости от места и способа получения исходного сырья, характера технологической обработки полупродуктов и т. п. Величины, приводимые по этому поводу в литературе (Ф. Е. Будагян, 1961; В. В. Ефремов, 1969; П. И. Шилов, Т. Н. Яковлев, 1964), характеризуют, как правило, уровень витамина до кулинарной обработки, которая сама по себе значительно разрушает тиамин. В среднем можно читать, что обычное приготовление пищи разрушает около 30% витамина. Некоторые виды обработки (высокая температура, повышенное давление и наличие больших количеств глюкозы), разрушают до 704-90% витамина, а консервация продуктов путем обработки их сульфитом может полностью инактивировать витамин В злаковых семенах других растений тиамин, подобно большинству водорастворимых витаминов, содержится в оболочке и зародыше. Переработка растительного сырья (удаление отрубей) всегда сопровождается резким снижением уровня витамина в полученном продукте. Шлифованный рис, например, совсем не содержит витамина.
Содержание тиамина в некоторых продуктах питания (В. В. Ефремов, 1960)
|
Продукт |
Содержание тиамина в мкг % |
Продукт |
Содержание тиамина в мкг % |
|
Пшеница |
0,45 |
Томаты |
0,06 |
|
Рожь |
0,41 |
Говядина |
0,10 |
|
Горох |
0,72 |
Баранина |
0,17 |
|
Фасоль |
0,54 |
Свинина |
0,25 |
|
Kpупa овсяная |
0,50 |
Телятина |
0,23 |
|
>>гречневая |
0,51 |
Ветчина |
0,96 |
|
>> манная |
0,10 |
Куры |
0,15 |
|
Рис шлифованный |
0,00 |
Яйца куриные |
0,16 |
|
Макароны |
следы |
Рыба свежая |
0,08 |
|
Мука пшеничная |
0,2—0,45 |
Молоко коровье |
0,05 |
|
>> ржаная |
0,33 |
Фрукты разные |
0,02—0,08 |
|
Хлеб пшеничный |
0,10—0,20 |
Дрожжи пивные сухие |
5,0 |
|
>>ржаной |
0,17 |
Орехи грецкие |
0,48 |
|
Картофель |
0,09 |
» земляные |
0,84 |
|
Капуста белокочанная |
0,08 |
4. Обмен тиамина в организме
Витамин поступает с пищей в свободном, эстерифицированном и частично связанном виде. Под влиянием пищеварительных ферментов происходит почти количественное его превращение в свободный тиамин, который всасывается из тонкого кишечника. Значительная часть поступившего в кровь тиамина быстро фосфорилируется в печени, часть его в виде свободного тиамина поступает в общий кровоток и распределяется по другим тканям, а часть снова выделяется в желудочнокишечный тракт вместе с желчью и экскретами пищеварительных желез, обеспечивая постоянную рециркуляцию витамина и постепенное равномерное усвоение его тканями. Почки активно экскретируют витамин в мочу. У взрослого человека за сутки выделяется от 100 до 600 мкг тиамина.. Введение повышенных количеств витамина с пищей или парентерально увеличивает выделение витамина с мочой, но по мере повышения доз пропорциональность постепенно исчезает и в моче наряду с тиамином начинают в возрастающих количествах появляться продукты его распада, которых при введении витамина свыше 10 мг на человека может быть до 40—50% исходной дозы (А. В. Труфанов, 1959). Опыты с меченым тиамином (А. Я. Розанов, 1960) показали, что наряду с неизмененным витамином в моче обнаруживается некоторое количество тиохрома, ТДС, пиримидиновый, тиалозовый компоненты и различные углерод- и серусодержащие осколки, в том числе меченые сульфаты.
Таким образом, разрушение тиамина в тканях животных и человека происходит достаточно интенсивно, но до настоящего времени не установлены реакции, через которые этот процесс осуществляется. Попытки обнаружить в животных тканях ферменты, специфически разрушающие тиамин, пока не дали убедительных результатов (см. ниже), хотя имеются отдельные сообщения по этому вопросу (Somogyi, 1966). Суммарное содержание тиамина во всем организме человека, нормально обеспеченного витамином, составляет примерно 30 мг, причем в цельной крови его находится 3—16 мкг%, а в других тканях (Williams е. а., 1950) значительно больше: в сердце — 360, печени — 220, в мозге — 160, легких — 150, почках — 280, мышцах — 120, надпочечнике — 160, желудке — 56, тонком кишечнике — 55, толстом кишечнике — 100, яичнике — 61, яичках — 80, коже — 52 мкг%. В плазме крови обнаруживается преимущественно свободный тиамин (0,1— 0,6 мкг%; Ю. М. Островский, 1957), а в эритроцитах (2,1 мкг на 1011 клеток) и лейкоцитах (340 мкг на 1011 клеток) — фосфорилированный.
Свободный тиамин в норме легко определяется в кишечнике и почках, что может быть связано и с недостатками чисто методического порядка, так как эти ткани обладают исключительно высокой фосфатазной активностью и к моменту взятия материала на исследование уже может происходить частичное дефосфорилирование эфиров витамина. С другой стороны, эти же механизмы могут играть определенную роль в удалении витамина из крови в мочу или кал. Количество витамина в последнем у человека составляет примерно 0,4—1 мкг и вряд ли связано в какой-то заметной степени с биосинтезом витамина кишечной микрофлорой.
Некоторое представление о динамике обмена тканевых запасов витамина дают опыты, проведенные S35-тиамином (Ю. М. Островский, 1971). Обновление тиамина происходит в разных тканях с различной скоростью и практически полная замена нерадиоактивного витамина на радиоактивный (вводимый ежедневно) осуществляется к 8-му дню опыта лишь в печени, почках, селезенке и скелетных мышцах. В сердце, поджелудочной железе ткани мозга к указанному сроку процесс этот не завершается. Вторая фаза опыта (авитаминозный режим) позволяет выявить ряд интересных закономерностей в отношении расходования эндогенных запасов витамина различными тканями. Немедленно и с наибольшей скоростью теряется метка тканью печени и поджелудочной железы. Равномерно с первого дня лишения животных меченого тиамина падает радиоактивность в селезенке, скелетных мышцах, почке. Практически на одном и том же уровне остается радиоактивность через сутки после лишения животных тиамина в ткани сердца и очень медленно теряется метка из мозга. По-видимому, в 1-й день авитаминозного режима миокард еще ассимилирует из крови необходимое количество тиамина, поступившего в нее туда из других органов. Уровень метки в крови (рис. 6) остается вначале нормальным и заметно падает лишь к 15-му дню опыта (7-й день авитаминоза).
Если в конце авитаминозного режима мышам снова начать вводить S35-тиамин, то никакого усиленного потребления метки тканями не наблюдается, т.е. предварительная девитаминизация не создавала условий, благоприятствующих (последующей ассимиляции витамина. Можно считать, что связывании витамина определяется, по-видимому, не дефицитом соответствующих депо, а какими-то лимитированными промежуточными реакциями специфической или неспецифической протеидизации витамина. Интерес представляет и некоторое замедление ассимиляции повторно вводимого витамина в ткани мозга, сердца и печени. Возможно, это обусловлено дефицитом соответствующих апоферментов или межтканевой конструкцией в захвате меченого тиамина из крови. Одновременно исследованная у тех же животных активность транскетолазы тканей только для крови коррелировала в какой-то мере со степенью девитаминизации всего организма. В других тканях между содержанием витамина и активностью фермента в длительные промежутки времени прямая связь не выявляется. Только в условиях, когда количество витамина уменьшается в несколько раз, такая связь начинает проявляться. Нечто аналогичное для соотношений между уровнем тиамина и активностью ДФ-содержащих ферментов недавно отмечено и другими авторами (Dreyfus, Hauser, 1965). Наиболее существенный вывод из представленных результатов состоит в утверждении, что количество витамина, находящегося в тканях, во много раз выше того уровня, который необходим для обеспечения специфических ферментных систем ТДФ. По-видимому, значительные количества витамина присутствуют в тканях, особенно в сердце и печени, в виде его производных, осуществляющих какие-то другие некоферментные функции.
4.1 Образование тиаминфосфатов (ТФ). За период с 1937 по 1943 г. показано, что фосфорилирование тиамина активно происходит в дрожжах и различных животных тканях. Уже тогда выяснилось, что реакция идет за счет АТФ по общему уравнению: тиамин + АТФ—> ТДФ + АМФ.
В 1952 г. эти закономерности были подтверждены на частично очищенном препарате тиаминкиназы из растворимой фракции гомогената печени. Оптимум рН для образования. ТДФ этим ферментным препаратом лежал, в пределах 6,8—6,9. Фосфорилирование тиамина подавлялось АМФ и АДФ. В присутствии АМФ образовывались лишь следы, а в присутствии АДФ — весьма незначительные количества ТДФ. Если в среду вместо тиамина вносился ТМФ, то образование ТДФ тормозилось. Очищенный примерно в 600 раз препарат тиамикиназы был применен (Forsander, 1956) для изучения Механизма фосфорилирования витамина с использованием меченой γ-Р32-АТФ. После выделения ТДФ Forsander пришел к выводу, что и вэтой системе тиамин получает от АТФ целиком пирофосфатную группировку.
Серия работ по изучению тиаминкиназы, выделенной из дрожжей и животных тканей, недавно проведена в Японии. На достаточно очищенных препаратах (более чем в 100 раз), обладавших слабой АТФ-азной активностью и не содержащих аденилаткиназы и нуклеозиддифосфокиназы, установлено, что ионы марганца, магния - кобальта активировали, а кальция, никеля, рубидия и железа — в широком диапазоне концентраций не угнетали фермент. На этом же препарате фермента показаны возможность фосфорилирования тиамина за счет других нуклеотидтрифосфатов (ГТФ, ИТФ, УТФ и др.) но то, что основным продуктом реакции является ТДФ и небольшое количество ТМФ. Применением Р32-АТФ, как и в исследованиях предыдущих авторов, подтвержден механизм переноса на тиамин сразу пирофосфатной группировки.
Однако результаты, полученные in vitro, не нашли полного подтверждения при изучении фосфорилирования тиамина на целых организмах и в опытах с митохондриями. С одной стороны, как будто подтверждалось предположение о пирофосфорилировании: после внутривенного введения тиамина уже через 30—60 минут в крови животных обнаруживались меченные по фосфору ТДФ и ТТФ, но не ТМФ. С другой стороны, после внутривенного введения ТМФ кокарбоксилазная и транскетолазная (Н. К. Лукашик, 1964) активность крови нарастала быстрее, чем после введения свободного тиамина. Некоторые микроорганизмы легче образуют ТДФ из ТМФ, чем из свободного витамина, а тиаминкиназа, найденная ранее в печени, не обнаружена в митохонд-риях почек, в которых фосфорилирование тиамина идет другим путем. Механизм фосфорилирования витамина с участием только АТФ не всегда укладывается в простую схему переноса пирофосфатной группировки в целом хотя бы потому, что наряду с ТДФ в различном биологическом материале обнаруживаются в значительных количествах и другие ТФ, в том числе даже Т-полифосфаты.
Ряд исследований последних лет касается вопроса о локализаций систем, ответственных за фосфорилирование тиамина. Печень уже через час после введения тиамина захватывает 33—40% витамина, накапливая различные его фосфорные эфиры. По данным А. А. Рыбиной (1959), происходит быстрое фосфорилирование меченого витамина и в других органах (в порядке убывающей активности): печень, почки, сердце, семенники, головной мозг. При этом радиоактивность фосфорных эфиров тиамина убывает в ряду: ТТФ, ТДФ, ТМФ. По данным японских авторов, фосфорилирование тиамина идет активно в митохондриях (Shima-zono, 1965), микросомах и гиалоплазме.
Из изложенных выше фактов нетрудно сделать вывод, что общая -интенсивность процессов эстерификции витамина в организме или в отдельных тканях должна в значительной степени коррелировать с активностью процессов, поставляющих АТФ. Первые экспериментальные наблюдения в этом плане, проведенные ня гомогенатах печени или клеточных элементах крови, получили в дальнейшем полное подтверждение. Все яды дыхания и гликолиза или соединения, конкурирующие с Т за АТФ, как правило, снижают уровень ТДФ в крови и в тканях.
4.2 Роль отдельных группировок в молекуле тиамина для его связывания в тканях.
За последние годы синтезировано более десятка новых производных тиамина (смешанные дисульфиды, О-бензольные производные и др.), широко внедряемых в лечебную и профилактическую практику. Преимущества новых витаминных препаратов, как правило, выявлялись чисто эмпирически в связи с тем, что до настоящего времени мы не располагаем достаточными сведениями о молекулярных механизмах ассимиляции тиамина, о характере его взаимодействия со специфическими (ферменты) и неспецифическими (осуществляющими транспорт витамина) белками. Необходимость точных представлений в этом вопросе диктуется и широкими перспективами использования антивитаминов тиамина (ампрол, хлоротиамин, деокситиамин) для лечебных целей (см. ниже).
Работы по синтезу новых производных тиамина с заранее заданными физико-химическими свойствами, обусловливающими возможности целенаправленного воздействия на обменные процессы в организме, немыслимы без конкретных представлений о роли отдельных групп атомов витамина и его производных в этой области. Значение пирофосфатного радикала для специфической потеидизации ТДФ в составе соответствующих ферментов уже отмечалось выше. В последние годы становится все более очевидным участие тиамина в других реакциях, не имеющих ничего общего с коферменными функциями витамина. Разнообразию активных группировок в молекуле тиамина соответствует каждый раз особая форма- претеидизации, блокирующая одни и обнажающая одновременно другие, важные для соответствующей функции, участки молекулы витамина (Ю. М. Островский, 1971). Действительно, первый тип протеидизации (через пирофосфатный радикал) отвечает коферментной функции и оставляет свободными, доступными для субстрата 2-й углерод тиазола и аминогруппу пиримидинового компонента. С другой стороны, очевидно, что участие витамина в окислительно-восстановительных реакциях или в процессах перефосфорилирования должно сочетаться с исключением возможности одновременного функционирования его как кофермента, так как в первом случае необходима деполяризация и раскрытие тиазолового цикла, а во втором — свободное положение фосфорилированного оксиэтильного радикала. Поскольку 80—90% тиамина, присутствующего в тканях, освобождается лишь при кислотном и ферментативном гидролизе, можно считать, что все связанные формы витамина находятся в протеидизированном, т. е. связанном с белками, состоянии.
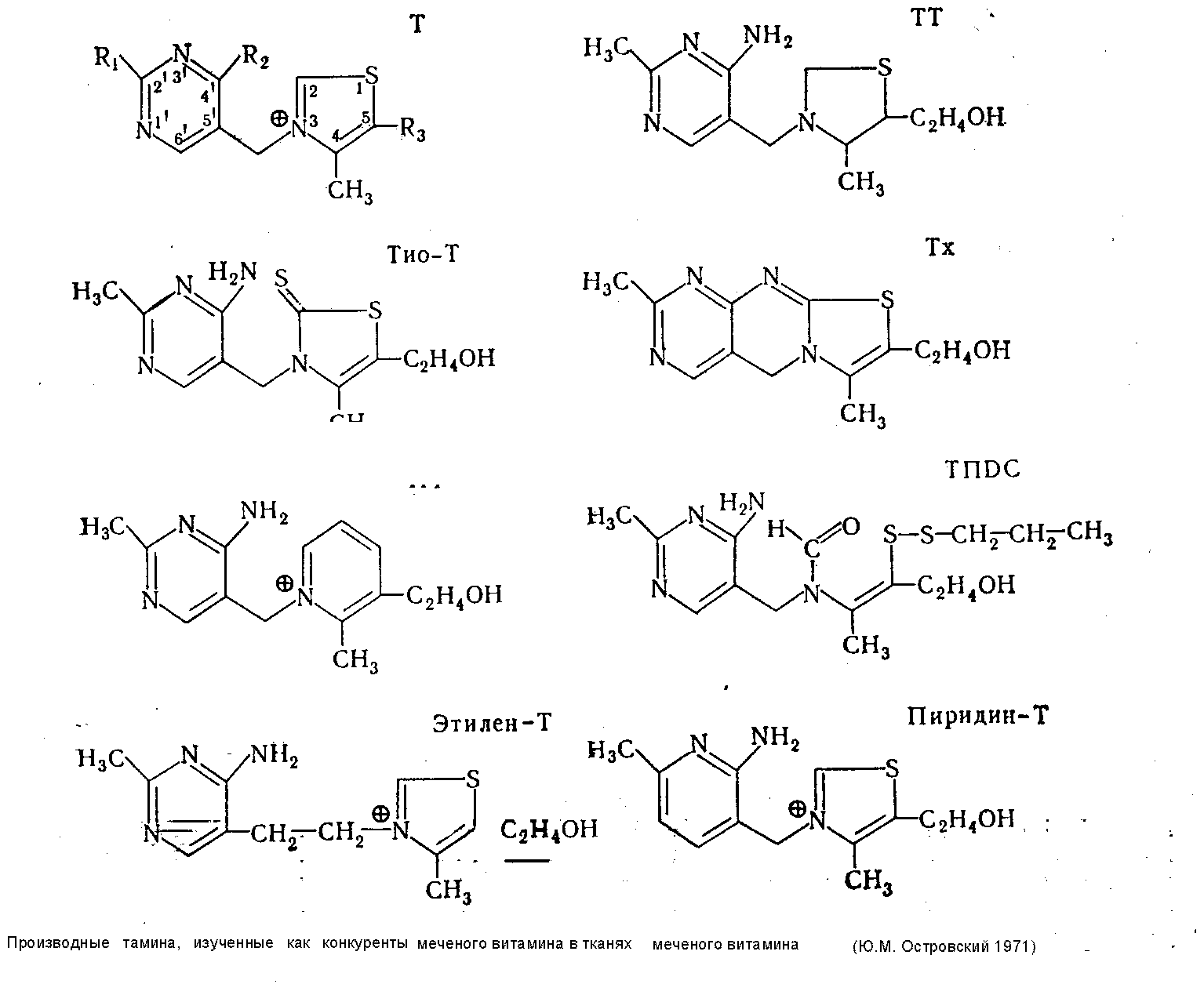
Представление о значении отдельных участков молекулы тиамина в этом процессе легко получить, определяя степень связывания тканями меченного по сере (S35) витамина и некоторых его производных, лишенных тех или иных активных центров, например аминогруппы — окситиамин (окси-Т), аминогруппы и оксиэтильного радикала — хлорокситиамин (ХОТ), четвертичного азота в тиазоловом цикле тетрагидротиамин (TТ) Некоторые ограничения на интерпретацию данных, полученных таким образом в короткие сроки опыта, накладывают физиологические механизмы, транспорта и экскреции вводимых соединений, в связи с чем дополнительно приходится исследовать также выведение витамина и самих меченых соединений с мочой. С другой стороны, результаты, получаемые в длительные сроки (24 часа), почти полностью соответствуют только представлениям о протеидизированной части витамина, а на основании рассмотрения конкурентных взаимоотношений между различными упомянутыми выше мечеными и другими немечеными производными витамина можно последовательно исключать роль отдельных атомов или группировок в механизмах фиксации тиамина тканями.
4.3 Коферментные функции тиаминдифосфата.
Известно значительное количество различных реакций, катализируемых ТДФ. Однако все их можно свести к нескольким типичным вариантам: простое и окислительное декарбоксидирование -кетокислот, ацилоиновая конденсация, фосфорокластическое расщепление кетосахаров. Ферментные системы, принимающие участие в этих реакциях по-видимому, едины в основных принципах своего действия; различна лишь последующая судьба «активного альдегидного осколка», возникающего на первых этапах процесса. Успехи, достигнутые в течение последних лет в изучении превращений -кетокислот (С. Е. Северин, 1964; А. А. Глемжа, 1964), позволили четко представить как роль собственно декарбоксилирующего фрагмента полиферментного комплекса дегидрогеназы, содержащего ТДФ, так и последовательность всех других, связанных с ним реакций:
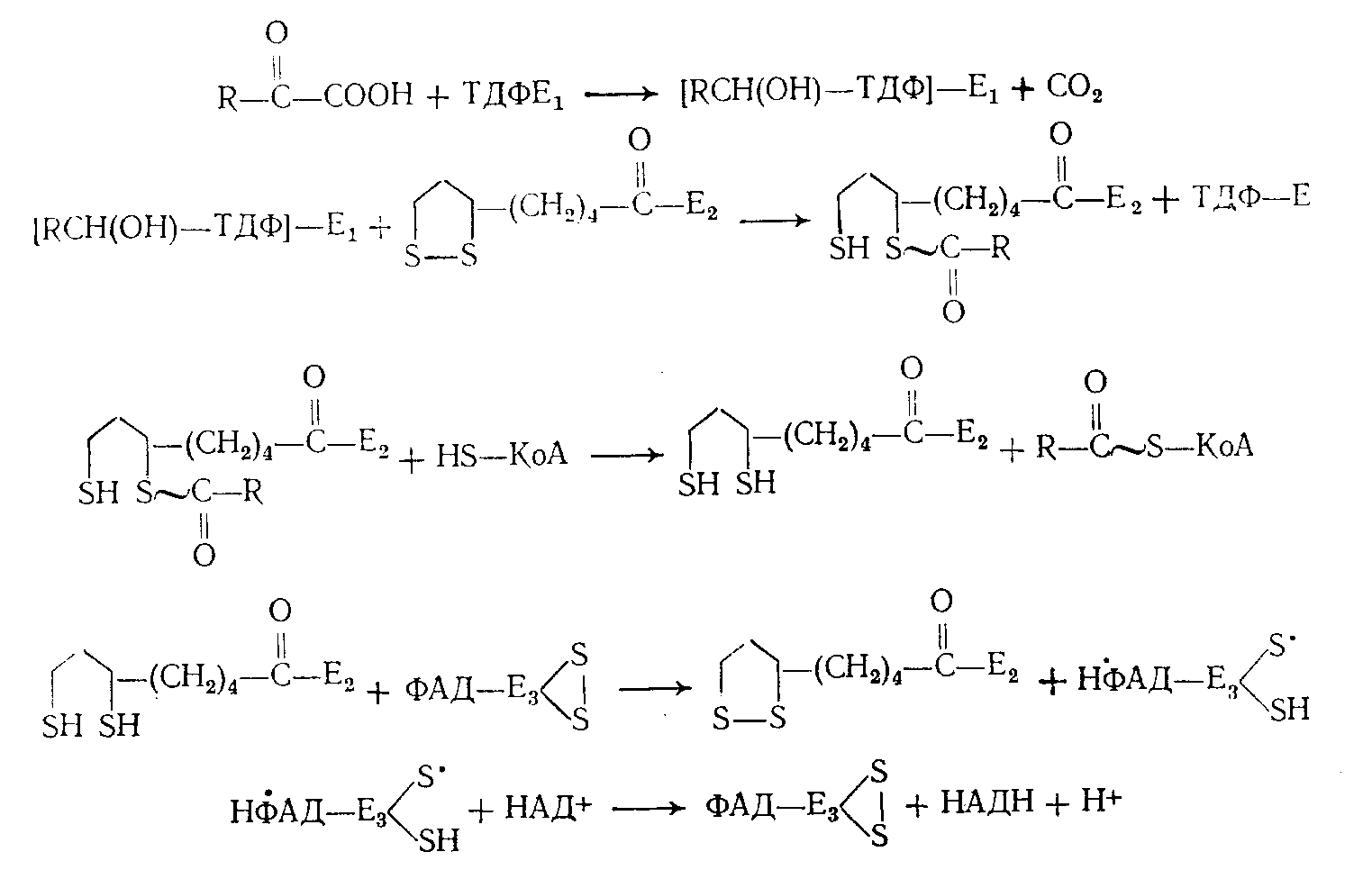
В системе транскетолазы (ТК) «активный альдегидный» осколок, очевидно, будет представлен гликолевым радикалом, переносимым от соответствующих источников (ксилулозо-5-фосфат, фруктозо-6-фосфат, оксипируват и др.) на различные акцепторы (рибозо-5-фосфат, эритро-зо-4-фосфат, глюкозо-6-фосфат). В фосфокетолазной реакции «активный гликолевый» радикал превращается непосредственно в ацетилфосфат.
Значительные успехи в выяснении механизма каталитического действия ТДФ были достигнуты в результате исследований, проведенных в двух основных направлениях: создание модельных неферментативных систем и введение в ферментные системы различных аналогов или антагонистов тиамина. Используя первый путь, удалось показать, что витамин и в нефосфорилированном виде способен при определенных условиях в отсутствие белка катализировать реакции декарбоксилирования, образования ацетона, дисмутации диацетила. Различными вариантами опытов, в которых коферментная активность ТДФ сравнивалась с активностью антиметаболитов витамина или изучалась с добавлением соли Рейнеке, бромацетата, пара-хлор-ртуть-бензоата и других соединений, показано, что каталитически наиболее важными группами в молекуле тиамина являются: сера, четвертичный азот тиазолового кольца, аминогруппа в положении 4 пиримидинового: кольца, второй углеродный атом тиазола (2-С-Тз), метиленовый мостик. Можно считать устоявшимся представление, что часть перечисленных выше активных центров (сера, азот, метиленовый мостик) крайне необходима только для поддержания определенной структуры и создания соответствующей электронной плот-ности-(Б. Пюльман, А. Пюльман, 1965) у второго углеродного атома тиазола (2-С-Тз), который является главным каталитическим центром. Спорными и неопределенными пока являются представления о значении аминогруппы пиримидинового компонента.
Некоферментная активность тиамина и некоторых его производных.
За последние 20 лет наряду выяснением механизма основных реакций, в которых каталитическую роль играет ТДФ, стали накапливаться данные о высокой биологической активности других некоферментных производных тиамина. Отчетливо наметились два направления исследований: возможное, участие различных фосфорных эфиров витамина в активном переносе богатых энергией фосфатных групп (ангидридная связь в ТДФ макроэргическая) и вероятность вмешательства тиамина в окислительно-восстановительные реакции В связи с тем что неизвестны специфические тиаминсодержащие ферментные системы, причастные к регуляции упомянутых выше процессов, наблюдаемые в этой сфере обмена эффекты витамина можно рассматривать как проявление его неспецифических функций.
Тиаминофосфаты (ТФ).
После разработки доступных методов получения ТДФ последний стали широко испытывать при различных заболеваниях в клинических условиях. Внутривенное введение 100—500 мг ТДФ при диабетическом ацидозе увеличивало количество пирувата, образующегося из глюкозы. Эффект аналогичного характера наблюдался при диабете после введения АТФ или фосфокреатина. Инкубация цельной крови с Р32н в присутствии Зх10-3М тиамина сопровождалась ускоренным внедрением метки в промежуточные продукты гликолиза (фруктозо-1,6-дифосфат, фруктозо-6-фосфат). Особенно заметно во времени, по сравнению с контролем без витамина, возрастала радиоактивность триозофосфатов. По данным Л. Н. Кузнецовой, Е. В. Лахно и Р. В. Чаговец (1953), в мышцах при утомлении и отдыхе распад и ресинтез ТДФ происходят примерно по тем же закономерностям, которые известны для АТФ и фосфокреатина. Характерными были изменения во время отдыха, когда количество ТДФ превышало исходный уровень до утомительной работы. Авторы не рассматривают причин усиленного распада ТДФ во время сократительной деятельности мышц, и вряд ли это возможно с позиции известных коферментных функций ТДФ. Торможение функциональной активности печени (введение ССЦ, гипоксия), по-видимому, за счет снижения расхода, сопровождается накоплением в органе ТФ. Наконец, совместное с глюкозой скармливание больших количеств ТДФ крысам сопровождалось заметным (на 69%) повышением уровня АТФ в печени.
Группа японских исследователей подробно изучила на циклофоразной системе внедрение Р32 из меченого ТДФ в немеченый АТФ. Перенос метки был в 3—4 раза больше в присутствии Р32-ТДФ, чем в присутствии неорганического Р32н, хотя система и во втором случае содержала столько же ТДФ. Если циклофоразную систему выделяли из печени страдающих от недостаточности тиамина крыс, то внедрение Р32 в АТФ из меченого ТДФ превышало контроль в 8—10 раз. Независимость внедрения Р32 в АТФ от эстерификации неорганического фосфата подтверждалась и тем, что ДНФ (10-4М), снижавший интенсивность окислительного фосфорилирования в 5—10 раз, не влиял на процесс переноса метки от ТДФ к АТФ.
Еще более интересными представляются результаты сравнительного изучения физико-химической и «субстратной» разнозначности концевых фосфатов АТФ и ТТФ. Оба соединения одинаково гидролизуются картофельной апиразой и кристаллическими препаратами миозина. Макроэргический характер фосфатных остатков в ТФ побудил ряд авторов исследовать роль этих соединений в начальных реакциях углеводного обмена. Пекарские дрожжи (Kiessling, 1957) в присутствии Р32н, и глюкозы интенсивно накапливают метку вначале преимущественно в -фосфате ТТФ и АТФ, а затем в α- и β-фосфатах. Удельная активность ТТФ в определенных условиях опыта может уравниваться с таковой для АТФ, что рассматривается как выражение причастности ТФ к переносу лабильных фосфатов при обмене глюкозы. Действительно, на белковых препаратах из дрожжей -Р32 ТТФ переносится на глюкозу в присутствии АДФ. Смесь ТФ (около 70% ТТФ), неактивная сама по себе, в присутствии АТФ стимулирует потребление глюкозы гексокиназой, полученной из миокарда, но не из скелетных мышц. Митохондрии содержат значительные количества ТФ, которые могут быть удалены на ѕ без заметного нарушения окисления ПК или сукцината. Сами эти ТФ захватывают до 75% Р32н по отношению к АТФ, а классический разобщающий яд ДНФ тормозит образование АТФ в большей степени, чем ТФ. Введение животным окси-Т, в зависимости от характера субстратов дыхания, может нарушать в митохондриях отдельно окислительные и фосфорилирующие реакции, а на интактных митохондриях, инкубируемых в отсутствие АТФ, добавление ТФ (особенно ТМФ) достоверно повышает эстерификацию неорганического фосфата. В последнее время установлено, что введение животным больших доз ТДФ уже через несколько часов значительно, (иногда в 2 раза) повышает в тканях содержание лабильных фосфорных соединений. Интересной представляется заметно большая эффективность ТМФ в ряде опытов, что, возможно, связано со своеобразным отношением этого производного тиамина к мембранам или обусловлено более легким его превращением в ТТФ. Есть основания полагать, что имеются определенные особенности взаимодействия ТФ с фосфорилирующими реакциями в различных тканях. В нервной ткани, например, с превращением ТФ ряд авторов связывают физиологические акты проведения возбуждения (Muralt, 1962) и транспорт ионов натрия через мембраны.
4.6 Свободный тиамин и его производные. Введение животным антиметаболитов витамина — окси-Т и ПТ — вызывает различную картину нарушений в обмене и в физиологических функциях, что позволило Д. Вулли (1954) предположить вероятность существования у тиамина нескольких различных или даже независимых друг от друга функций. Различие между этими антиметаболитами с химической точки зрения сводится к исключению тиолдисульфидных превращений у ПТ и три-циклических по типу тиохрома (Тх) у окси-Т. Возможность каталитического действия тиамина на уровне окислительно-восстановительных реакций в обмене давно допускают и критикуют разные авторы. Действительно, различная обеспеченность витамином сильно влияет на активность ряда окислительных ферментов или содержание в крови восстановительных форм глютатиона. Витамин обладает антиоксидантными свойствами в отношении аскорбиновой кислоты, пиридоксина и легко взаимодействует с оксигруппами полифенолов (Takenouchi, 1965). Дигидро-Т частично окисляется в тиамин дрожжами и бесклеточными экстрактами, кристаллическими препаратами пероксидазы, тирозиназы и неферментативно при взаимодействии с кристаллическим убихиноном, пластохиноном, менадионом.
5. Физиологическое значение витамина В1 состоит в мощном регулирующем воздействии его на отдельные функции организма и, в первую очередь, на обменные процессы. Сущность же этого воздействия заключается в том, что тиамин участвует в обмене веществ в качестве коэнзима. Особенно важную роль витамин В1 играет в углеводном обмене.
Свою биологическую активность тиамин приобретает в кишечнике, печени и почках в процессе присоединения фосфорной кислоты (фосфорилирования).
Значение витамина В1 для нормального обмена углеводов и состоит прежде всего в том, что фосфорное производное тиамина – дифосфотиамин (кокарбоксилаза) в виде простетической группы входит в состав карбоксилазы – фермента, который с участием липоевой кислоты осуществляет реакции декарбоксилирования (отщепление СО2) пировиноградной и других α-кетокислот с последующим их распадом.
Если в организме мало тиамина, то дальнейший распад пировиноградной кислоты задерживается. Это подтверждается повышением уровня пировиноградной кислоты в условиях В1-витаминой недостаточности. Наряду с этим показано, что уровень пировиноградной кислоты может повышаться и при других патологических процессах, сопровождающихся нарушением углеводного обмена, как, например при болезни Боткина, сахаром диабете и др. Витамин В1 снижает уроень пировиноградной кислоты и у здоровых людей, причем такой же эффект выявлен и после предварительной нагрузки глюкозой. Проведено также изучение влияния витамина В1 на процессы усвоения сахара мышцами в патологических условиях. По данным исследования артерио-венозной разницы в содержании сахара при двойной сахарной нагрузке до и после введения витамина В1 (20мл) выявлено положительное значение этого витамина для регуляции нарушенных процессов углеводного обмена (З.В. Новицкая, 1950).
Особого внимания заслуживает значение витамина В1 для функционального состояния центральной нервной системы и мышечной деятельности. Это становится понятным, если учесть, что центральная нервная система почти всю свою энергию черпает из углеводов, в обмене которых тиамин принимает активное участие. Нарушение В1 – витаминного баланса лишает центральную нервную систему возможности эффективно использовать глюкозу при одновременном накоплении здесь промежуточных продуктов обмена (пировиноградная, молочная кислота и др.) токсически влияющих на мозг.
Тиамин является важным фактором в передаче нервных импульсов – тормозит образование и инактивирует холинэзтеразу, которая гидролизирует ацетилхолин. Этим самым тиамин косвенно усиливает активность ацетилхолина как передатчика нервного возбуждения.
Последующие исследования установили, что В1 – авитаминоз (полиневрит) всегда сопровождается глубокими нарушениями белкового обмена в силу уменьшения активности различных ферментов, принимающих участие в этом обмене.
Особого внимания заслуживает участие тиамина в синтезе нуклеиновых кислот. Дефицит его так же приводит к нарушению гемопоэза.
Таким образом, можно прийти к общему выводу, что витамин В1, являясь мощным регулятором углеводного обмена, в то же время имеет важное значение и для обмена белков. За последнее время получены отчетливые данные о связи этого витамина также с жировым, минеральным и водным обменом.
Кроме того, тиамин оказывает так же выраженное влияние на функциональное состояние отдельных органов и систем организма. Связанные с гипо- и авитаминозом нарушения центральной нервной системы, в свою очередь, ведут к расстройству сердечно-сосудистой системы, эвакуарно-секретной функции желудка и кишечника и эндокринной системы.
Тиамин обладает относительно низкой фармокологической активностью. Все приведенные факты заставляют думать, что описанные для витамина многочисленные положительные эффекты разрешаются каким-то образом на более специфически связанных с ним обменных уровнях или в системах, которые пока ускользают от контроля.
План:
Биотин…………………………………………………………..1
Исторические сведения……………………………………..1
Химические и физические свойства биотина………………1-3
Аналоги и ингибиторы……………………………………………3-4
Распространение биотина в природе…………………………..5
Биосинтез биотина……………………………………… 5-7
Обмен биотина в организме…………………………………….7-9
Участие биотина в обмене веществ и механизм действия………………………………………………………9-12
Взаимодействие с другими витаминами………12-13
Потребность организма в биотине……..…………….13
Проявление недостаточности биотина………..13-14
8. Профилактическое и лечебное применение биотина….14-15
Тиамин…………………………………………………………..16
Исторические сведения……………………………….16
Химические и физические свойства витамина …16-17
Распространение витамина В1, в природе………..18
Обмен тиамина в организме………………………18-20
Образование тиаминфосфатов (ТФ)..………..20-21
Роль отдельных группировок в молекуле тиамина для его связывания в тканях………………….21-22
Коферментные функции тиаминдифосфата…22-23
Некоферментная активность тиамина и некоторых его производных……………………………..………23
Тиаминофосфаты (ТФ)………………………...…23-24
Свободный тиамин и его производные..………24-25
5. Физиологическое значение.…………………………25-26
Список использованной литературы
Витамины. Каталог. М., 1957
Крыжановская И.И. Витамин В1 при хронической недостаточности кровообращения. Киев 1967.
Виноградов В.В. Гормональные механизмы метаболического действия тиамина Минск. 1984.
Островский Ю.М. Активные центры и группировки в молекуле тиамина. Минск. 1975.
Познанская А.А. Витамины М. 1974.
Биохимия и физиология витаминов под ред. Проф. М.М. Сисакина М. 1950.
Тиамин. Обмен, механизм действия (сборник статей) отв. Ред. А.А. Титаев. М. 1978.
Островский Ю.М. Тиамин. Минск 1971.
Министерство здравоохранения Р.Ф.
Ярославская Государственная Медицинская Академия
Кафедра биологической и биоорганической химии
Р Е Ф Е Р А Т
Тема: «Строение, свойства и биологическая роль биотина и тиамина.»
Выполнила:
Студентка I-го курса
13-ой группы, пед. Факультета
Бородулиной Е.А.
Руководитель:
Хохлова О.Б.
Ярославль 1999