
Рефераты по рекламе
Рефераты по физике
Рефераты по философии
Рефераты по финансам
Рефераты по химии
Рефераты по хозяйственному праву
Рефераты по цифровым устройствам
Рефераты по экологическому праву
Рефераты по экономико-математическому моделированию
Рефераты по экономической географии
Рефераты по экономической теории
Рефераты по этике
Рефераты по юриспруденции
Рефераты по языковедению
Рефераты по юридическим наукам
Рефераты по истории
Рефераты по компьютерным наукам
Рефераты по медицинским наукам
Рефераты по финансовым наукам
Рефераты по управленческим наукам
Психология и педагогика
Промышленность производство
Биология и химия
Языкознание филология
Издательское дело и полиграфия
Рефераты по краеведению и этнографии
Рефераты по религии и мифологии
Рефераты по медицине
Рефераты по сексологии
Рефераты по информатике программированию
Краткое содержание произведений
Реферат: Гемоглобин эритроцитарных мембран человека
Реферат: Гемоглобин эритроцитарных мембран человека
содержание
1. ВВЕДЕНИЕ. 3
2. ОБЗОР ЛИТЕРАТУРЫ.. 6
3. МАТЕРИАЛ И МЕТОДЫ ИССЛЕДОВАНИЯ
3.1. Материал исследования. 29
3.2. Методы исследования. 29
4. РЕЗУЛЬТАТЫ СОБСТВЕННЫХ ИССЛЕДОВАНИЙ
4.1. Количественное содержание гемоглобина и основных белков мембран эритроцитов человека. 34
4.2. Взаимное варьирование количественной представительности отдельных белков эритроцитов человека. 36
4.3. Обсуждение. 39
5. ЗАКЛЮЧЕНИЕ. 40
6. ВЫВОДЫ.. 41
7. СПИСОК ЛИТЕРАТУРЫ.. 42
Введение
Мембрана эритроцита в течение долгого времени представлялась исследователям лишь оболочкой, отделяющей гемоглобин от плазмы. Роль мембраны в функционировании этой высоко специализированной клетки, казалось, ограничивается только способностью быть проницаемой для газов крови. Однако успехи, достигнутые в мембранологии за последние годы и, в частности, изучении мембраны эритроцита млекопитающих с помощью биохимических и биофизических методов, заставляют в настоящее время по-иному взглянуть на роль мембраны в работе клетки. Совокупность имеющихся в литературе данных позволяет сделать вывод о том, что плазматическая мембрана эритроцита – важнейший элемент клетки; она одновременно является и механической оболочкой с регулируемыми физическими свойствами, и "диспетчерской" клетки, осуществляющей координацию работы клетки в зависимости от физических и химических сигналов, поступающих к ней в организме. [9]
Важнейшие компоненты биологических мембран – белки, которые осуществляют их специфические функции. Одни из них обеспечивают транспорт определенных молекул внутрь клетки или из нее, другие являются ферментами и катализируют ассоциированные с мембраной реакции, третьи осуществляют структурную связь цитоскелета с внеклеточным матриксом или служат в качестве рецепторов для получения и преобразования химических сигналов из окружающей среды. Мембранные белки, образующие цитоскелет, соединяются друг с другом специфическим для каждого типа клеток образом и формируют сложные динамически взаимодействующие между собой структуры. Эти структуры и отвечают непосредственно за согласованное поведение частей клеток. [4] Гемоглобин можно считать своего рода модельным белком, структура, свойства и функции которого наиболее полно изучены по сравнению с другими белками на протяжении последних 50 лет. Десятки лет исследований гемоглобина во многих лабораториях мира привели к значительному прогрессу в описании и понимании физических, химических и биологических аспектов его функционирования. [2] Однако, несмотря на это, в научной литературе недостаточно подробно освещен вопрос о связи гемоглобина со структурными белками эритроцитарных мембран.
Изучение структуры мембраны эритроцита человека и прежде всего входящих в их состав белков является актуальным, поскольку ряд наследственных патологий (сфероцитарные анемии, миодистрофии, некоторые заболевания центральной нервной системы) характеризуются различными нарушениями в белковой компоненте эритроцитарной мембраны и как следствие нарушением ее структурно-функциональной организации. [7] Определенные факторы среды могут изменять экспрессию генов, что отражается на жизнедеятельности как отдельных органов и систем, так и организма в целом. Некоторые авторы рассматривают уровень гемоглобина в крови как универсальный неспецифический показатель адаптационных процессов, процессов напряженности организма в ответ на различные внешние воздействия. [1] Так, следствием действия таких факторов может явиться уменьшения количества гемоглобина в крови или нарушения структурно-функциональной организации белковой компоненты мембран эритроцитов. Результатом таких изменений служит ряд патологий, и, как следствие, - значительное снижение адаптационных возможностей человека. Поэтому на сегодняшний день актуально изучение структуры мембрансвязанного гемоглобина, оценка его количественного содержания и взаимосвязь со структурными белками эритроцитарных мембран. В связи с начавшимся систематическим изучением продуктов экспрессии генов человека в рамках молекулярно-анатомического направления представляется крайне важным установление количества и основных свойств полипептидов, входящих в состав мембраны эритроцита, и составление каталога белков. Возможности этих исследований, а также поиска первичного биохимического дефекта при наследственных аномалиях существенно возросли после широкого распространения метода двухмерного электрофореза в полиакриламидном геле (ПААГ). [7]
Целью настоящего исследования явилось изучение количественного содержания мембраносвязанного гемоглобина эритроцитов человека и его связи со структурными белками.
литературный обзор
В силу своей специализации (перенос кислорода от легких к тканям) эритроцит по механическим свойствам является уникальной клеткой, так как в отличие от других клеток постоянно подвергается выраженным деформирующим воздействиям в кровеносном русле. [9]
Для нормального функционирования эритроцит должен в течение относительно длительного периода времени сохранять свою целостность и быть хорошо деформируемым; последнее свойство важно прежде всего для нормальной микроциркуляции. В свою очередь деформируемость определяется рядом факторов, основные из которых – форма клетки и эластичность мембраны. Сейчас можно считать доказанным, что эти свойства мембраны эритроцита и клетки в целом обусловлены наличием белковой мембранной структуры, именуемой мембранным скелетом клетки и расположенной на внутренней стороне мембраны. [9] Плазматическую мембрану эритроцитов следует рассматривать как открытую динамическую структуру, способную специфически и неспецифически связывать белки плазмы крови и цитозоля эритроцитов. [6] Исследователи пытаются выяснить механизмы метаболического регулирования белковых взаимодействий компонентов мембранного скелета между собой, а также с интегральными белками мембраны, что должно подвести к пониманию природы таких процессов, как изменение формы и деформируемости, работа транспортных систем, изменение ионной проницаемости мембраны. В данном обзоре сделана попытка обобщить и осмыслить имеющиеся данные по этому вопросу.
Современные представления о структуре и функции мембранного скелета эритроцитов человека и других млекопитающих сложились в основном за последние 25 – 30 лет. За этот период опубликован ряд подробных обзоров. Наличие сетчатой структуры, выстилающей внутреннюю поверхность мембраны эритроцита, было обнаружено непосредственно с помощью электронной микроскопии после обработки клеток неионным детергентом тритоном Х-100. Мембранный скелет виден в виде двухмерной сети филаментов, длина которых зависит от особенностей приготовления препарата для микроскопии. Белковые компоненты мембранного скелета были идентифицированы с помощью электрофореза в полиакриламидном геле (ПААГ) в присутствии додецилсульфата натрия (ДДС).
Классической работой по электрофоретическому разделению белков мембраны эритроцитов человека является работа Фейрбанкса и соавторов, в которой авторы предложили номенклатуру полипептидных полос, выявляемых в солюбилизированных с помощью ДДС мембране; этой номенклатурой пользуются в настоящее время большинство исследователей. При одновременном электрофорезе солюбилизированных белков эритроцитарной мембраны в ПААГ в присутствии ДДС выявляется 20 полос, однако Фейрбанкс с соавторами выделили 7 основных полос, которые были пронумерованы от катода к аноду. В последующие годы были внесены лишь незначительные уточнения в эту номенклатуру. [9]
В связи со способом расположения макромолекул в мембране эритроцита белки делят на периферические (внутренние) и интегральные. Интегральные удерживаются в мембране. В их состав входят гликопротеины и протеолипиды. Функции интегральных белков разнообразны: они могут выступать в роли гидролитических ферментов, рецепторов клеточной поверхности, окислительно-восстановительных компонентов транспортной системы электронов и в качестве специфических белков входит в состав цитоскелета, который представляет собой двумерную сеть, соединенную непосредственно с мембраной эритроцита через взаимодействие с интегральными белками. Также к периферическим белкам относится ряд эритроцитарных ферментов.
Метод фракционирования мембранных белков одномерным электрофорезом в ПААГ в присутствии ДДС позволили разделить белки мембраны относительно их электрофоретической подвижности, и полученным фракциям белков были даны номера по мере уменьшения их молекулярной массы.
Полосы 1 и 2 Включают отдельные a- и b-субъединицы спектрина.
Спектрин – основной белок цитоскелета эритроцитов. Спектрин образует двумерную сеть, к которой крепятся актиновые олигомеры. Молекула спектрина представляет собой ab-гетеродимер. По данным электронной микроскопии, гетеродимеры имеют форму изогнутых фибрилл длиной около 100 нм. Они могут принимать разные конфигурации. Каждая цепь молекулы спектрина содержит более 2000 аминокислотных остатков и состоит из линейно расположенных структурных доменов. Доменная структура была установлена с помощью анализа пептидных фрагментов субъединиц. Вторичная структура a- и b-субъединиц сходна и представляет собой a-спиральные участки, разделенные чувствительными к протеолизу b-структурами. Димеры спектрина ассоциируются с b-цепью другого димера "голова к голове". Никогда не встречаются a-a- или b-b-связи. Между димерами и тетрамерами существуют обратимые переходы, которые зависят от условий среды. При 37 °С спектрин экстрагируется из мембран эритроцитов в форме димера, при 4 °С – в форме тетрамера. При 30 и 40 °С в растворе между димерами и тетрамерами устанавливается равновесие. Так как в обычных условиях получения спектрин экстрагируется в форме тетрамера, полагали, что in vivo спектрин находится в той же форме. Электронно-микроскопический анализ свидетельствует о том, что такие олигомеры образуются в результате ассоциации 3-7 димеров спектрина. Полагают, что в эритроцитах присутствуют как олигомеры, так и тетрамеры, которые образуют двумерную сеть цитоскелета. Молекула спектрина подвергается множественному фосфорилированию, степень которого не влияет на форму эритроцитов. [5]
Полосы 2.1, 2.2, 2.3 Представлены различными формами периферического белка анкирина. Данные электронной микроскопии свидетельствуют о том, что сеть спектрина расположена на определенном расстоянии от мембраны и, по-видимому, взаимодействует интегральными белками мембраны. Латеральная подвижность интегральных белков зависит от присутствия спектрина: это подтверждает наличие такого взаимодействия. При обработке мембран химотрипсином был выделен белковый фрагмент с мол. массой 72 kD, который ингибировал взаимодействие спектрина с эритроцитарной мембраной, обедненной спектрином. Белок был назван анкирином, от греческого ankyra, что значит заякоривать. Другая группа исследователей назвала его синдеином, от греческого syndeo, что значит связываться вместе. Молекула анкирина (мол. масса 200 kD) имеет сферическую форму и состоит из двух доменов: нейтрального и фосфорилированного. Фосфорилированным доменом анкирин взаимодействует с b-субъединицей спектрина на расстоянии 20 нм от С-конца молекулы. Нейтральный домен анкирина осуществляет связь с белком полосы 3. [5,6]
Белок полосы 3 Белок 3 (анионтранспортный белок) – один из основных интегральных белков эритроцитарной мембраны – является гликопротеидом с мол. массой 90 kD. Показано, что этот белок принимает участие в обеспечении анионного транспорта, в связывании цитоскелетного каркаса с мембраной, цитоплазматических белков гемоглобина и глицеральдегид-3-фосфатдегидрогеназы. Не все молекулы белка 3 взаимодействуют с анкирином. Если бы все молекулы белка 3 были связаны с анкирином и, следовательно, со всей спектриновой сетью, это исключило бы возможность латеральных перемещений самого белка, образования кластеров и транспортных функций. Связь спектрина с мембраной осуществляется, вероятно, не только через анкирин, но также с помощью дополнительных взаимодействий. Имеющиеся на сегодняшний день данные свидетельствуют о том, что белок 4.1 обеспечивает связь спектрина с интегральным белком мембраны. [5] Все перечисленные дополнительные взаимодействия носят слабый характер и, видимо, играют второстепенную роль по сравнению со взаимодействием спектрин―анкирин―белок 3.
Белок полосы 4.1 Необходим для нормальной стабилизации мембраны, закрепляет связь спектрина с актином. На электрофореграмме данный белок разделяется на две полосы: 4.1а (Mr=80 kD) и 4.1b (Mr=78 kD). Переход 4.1а в 4.1b осуществляется с помощью деамидирования. Это глобулярный белок с мол. массой 80 kD, составляющий около 5% от белковой массы цитоскелета эритроцитов. Его изоформы сходны по аминокислотному составу. Обе изоформы способны взаимодействовать со спектрином. Каждая из них содержится в количестве примерно 100 000 молекул на 1 клетку. [5]
Белок полосы 4.2 Паллидин – это олигомер (Mr=72 kD), в клетке присутствует в димерной и тримерной формах с Mr=185 kD. Соединяется с цитоплазматическим доменом белка полосы 3, анкирином, спектрином и белком полосы 4.1. [14]
Белок полосы 4.5 В его состав входят интегральные белки – переносчики нуклеозидов и глюкозы через эритроцитарную мембрану. Основная часть белков полосы 4.5 представлена транспортерами глюкозы (Mr=55 kD). [14]
Белок полосы 4.9 Центральный компонент цитоскелета с Mr=48 kD. Этот белок выделен в виде тримера с Mr=145 kD. Стабилизирует взаимодействия спектрина с актином и влияет на степень его полимеризации. Это фосфопротеин. Белок присутствует в эритроцитах в количествах, приблизительно равных количеству спектрина. О функциях этого белка практически ничего не известно. Возможно, он принимает участие в укреплении цитоскелетного каркаса (сети цитоскелета). Показано, что очищенный полипептид полосы 4.9 может взаимодействовать с актиновыми нитями, снижая скорость полимеризации актина и, возможно, стабилизируя короткие актиновые нити в эритроцитах. [5]
Полоса 5 Актин – представлен в эритроцитах b-изоформой и по физико-химическим свойствам схож с актином поперечнополосатых мышц. Так как функциональной формой актина является его полимер, изначально пытались выяснить, есть ли в эритроцитах филаментный актин. Электронно-микроскопические исследования показали, что актин присутствует в эритроцитах в виде филаментов, содержащих от 12 до 17 мономеров актина. [5] Выделенные мембраны эритроцитов, а также комплекс спектрина, актина и белка 4.1 способны индуцировать полимеризацию мономерного актина, подобно олигомерам актина. Обработка цитохалазином В подавляет полимеризацию. Так как типичный фибриллярный актин не выявляется в нативных эритроцитах, а в опытах in virto можно индуцировать образование длинных филаментов, связанных с мембраной, следует, по-видимому, предположить, что в эритроцитах существуют какие-то механизмы, ограничивающие рост филаментов, подобно тому, как это делает цитохалазин В. Актиновые олигомеры связываются с N-концом молекулы спектрина латерально. Каждый тетрамер спектрина имеет два сайта связывания для актиновых олигомеров. Спектрин может связываться с Ф-актином по всей длине актиновой нити, и эти связи, видимо, существенны для образования цитоскелетной сети. Важная роль во взаимодействии актина и спектрина принадлежит белку полосы 4.1, который связывается со спектрином в тех же участках, что и актин. Связь спектрина с актином в отсутствие белка 4.1 слабая и значительно усиливается в его присутствии, поэтому взаимодействие спектрина и актина называют 4.1-зависимым. Комплексы белка 4.1, спектрина и актина обладают гораздо большей устойчивостью к диссоциации при седиментации в сахарозе, чем комплексы актина и спектрина.
Возникает вопрос о том, какие факторы оказывают влияние на полимеризацию эритроцитарного актина. Относительно низкая концентрация актина в эритроцитах не может, вероятно, иметь решающего влияния на длину филаментов. Предположим, что на полимеризацию актина могут влиять такие актинсвязывающие белки, как спектрин и белок 4.1. Оказалось, что эти белки способны регулировать скорость полимеризации актина, сокращая лаг-фазу, уменьшая скорость элонгации и не вызывая при этом никаких изменений в последовательности этапов полимеризации. Спектрин и белок 4.1 вызывают нуклеацию глобулярного актина в условиях, когда концентрация актина ниже критической и спонтанная полимеризация не происходит. Таким образом, можно предположить, что спектрин и белок 4.1 представляют собой комплекс белков, блокирующих медленный конец актиновой нити в отличие от большинства "запирающих" белков, взаимодействующих с быстрым концом. Однако позже были получены данные, которые противоречат этому предположению. Шен с соавторами выделили фрагменты цитоскелета эритроцитов при обработке растворами с низкой ионной силой при 37 °С. [5] Электронно-микроскопические исследования полученных фрагментов показали, что часть из них содержит филаменты актина, вдоль которых кластерами располагаются спектриновые молекулы. При инкубации этих препаратов с глобулярным актином (при концентрации актина в растворе выше критической) происходил рост актиновых нитей на обоих концах. Тсукита с соавторами изучали полимеризацию актина на мембранах эритроцитов, закрепленных на электронно-микроскопических сектах. При инкубации этих препаратов с глобулярным актином на внутренней поверхности мембран, содержащих цитоскелетные белки наблюдался рост актиновых филаментов. [5] При концентрациях глобулярного актина, близких к критическим, рост наблюдался только на медленном конце, а при дальнейшем увеличении концентрации – на обоих концах нити. При обработке мембран "кэпирующим" белком с мол. массой 45 kD, связывающимся с быстрым концом актиновых микрофиламентов, рост наблюдался только на медленном конце. Данные Шена и Тсукиты приводят к выводу о том, что в эритроцитах актин существует в виде филаментов со свободными концами. Следовательно, спектрин и белок 4.1 не являются кэпирующими белками, однако несомненно влияют на состояние актина в эритроцитах, стабилизируя актиновые филаменты.
Белок полосы 6 Глицеральдегид-3-фосфатдегидрогеназа (ГАФД), молекулярная масса составляет 35 kD. Располагается на цитоплазматическом домене анионтранспортного белка, в составе мультиферментного комплекса. Данный белок является ферментом гликолиза и принимает участие в регуляции окисления гемоглобина. [14]
Полоса 7 Основной составляющей полипептид – тропомиозин. Долгое время оставалась неизвестной природа белка зоны 7. Было показано, что этот белок связывается с антителами к тропомиозину, выделенному из мускульного желудка цыпленка. Молекулярная масса белка совпадала с молекулярной массой тропомиозинов, выделенных из мозговых тканей, из макрофагов и кровяных пластинок. Тропомиозин эритроцитов – димер с мол. массой 60 kD, состоящий из двух субъединиц с мол. массой 29 и 27 kD и связывающийся с мышечным актином (Ф-актином) при соотношении: 1 молекула тропомиозина на 6-7 мономеров актина. Так как в эритроцитах филаменты актина содержат до 15-17 мономеров, они могут связывать две молекулы тропомиозина. [5]
Полоса 8 Представлена пептидной частью фермента глутатион-S-трансферазы (Mr=23 kD). Взаимосвязь белка полосы 8 с мембраной является кальций – зависимой.
В 1985 г. было показано, что еще одним компонентом цитоскелета эритроцитов является миозин. С помощью моноклональных антител к тяжелой цепи миозина Фаулер обнаружил в эритроцитах полипептид, который был выделен и охарактеризован. Белок состоит из тяжелой цепи с мол. массой 200 kD и из двух легких цепей с мол. массами 26 и 19,5 kD. В растворах с низкой ионной силой он образует биполярные филаменты и обладает АТФазной активностью. В эритроцитах миозин присутствует примерно в количестве 6000 молекул на 1 клетку, т.е. на 1 молекулу миозина, как и в других клетках, приходится около 80 мономеров актина. [5]
Белки, образующие цитоскелет эритроцитов, неуникальны. Они или им подобные белки есть во многих других клетках. Преимущественная их локализация – вблизи мембран. Вероятно также, что рассмотренные белки могут быть полифункциональны и использоваться в структурных комплексах разного функционального назначения. [5]
Полосу 9 на электрофореграмме составляет белок глобин, который является частью мембрансвязанного гемоглобина. Как уже говорилось выше, цитоплазматический домен белка полосы 3, включающий около 380 аминокислотных остатков, локализован в цитоплазме и играет важную роль в поддержании формы эритроцита и его метаболизме. N-концевая последовательность цитоплазматического домена богата аминокислотными остатками кислого характера. На этом участке находятся центры связывания гидролитических ферментов – глицеральдегид-3-фосфатдегидрогеназы, альдолазы, каталазы, гемоглобина и гемихрома. Таким образом, цитоплазматический домен анионтранспортного белка и является местом локализации мембрансвязанного гемоглобина, о структуре, свойствах и некоторых аспектах функционирования которого будет сказано ниже.
Гемоглобин Гемоглобин – белок эритроцитов, красных кровяных клеток, переносящий молекулярный кислород от легких к тканям в организмах позвоночных животных. Гемоглобин можно считать своего рода модельным белком, структура, свойства и функции которого наиболее полно изучены по сравнению с другими белками на протяжении последних 50 лет. Американский физик Хопфилд назвал его атомом водорода современной биохимии, имея в виду, что изучение гемоглобина сыграло в биохимии ту же роль, что и изучение атома водорода в физике. Гемоглобин называют также почетным ферментом, поскольку исследования его структуры в статике и динамике позволили значительно продвинуться в понимании механизмов функционирования ферментов. Структура этого глобулярного белка известна в деталях главным образом благодаря работам английского биофизика Макса Перутца, который получил первые рентгеноструктурные данные еще в конце 40-х годов нашего века. За эти исследования он был удостоен Нобелевской премии.
Структура гемоглобина Молекула гемоглобина состоит из четырех субъединиц: двух a и двух b - и соответственно содержит четыре полипептидные цепочки двух сортов. Каждая a-цепочка содержит 141, а b-цепочка – 146 аминокислотных остатков. Таким образом, вся молекула гемоглобина включает 574 аминокислоты. Хотя аминокислотные последовательности a- и b-цепочек различны, они имеют практически одинаковые третичные пространственные структуры. Собственно говоря, приведенные выше детали структуры относятся не к гемоглобину, а к его белковой компоненте – глобину. Каждая субъединица гемоглобина содержит одну небелковую (так назваемую простетическую) группу – гем. Гем представляет собой комплекс Fe(II) с протопорфирином. Структура гема представлена на рис.1, а. [2]
Атом железа может образовать шесть координационных связей. Четыре связи направлены к атомам азота пиррольных колец, оставшиеся две связи –
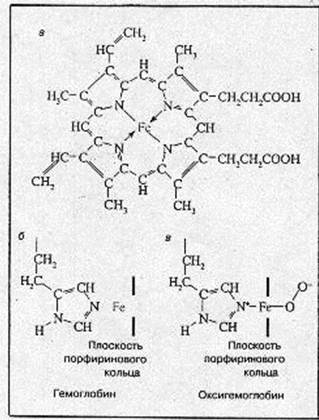
Рис.1. Структура гема (а), структура активного центра дезоксигемоглобина (б), структура активного центра оксигемоглобина (в).
перпендикулярно к плоскости порфиринового кольца по обе его стороны. Гемы расположены вблизи поверхности белковой глобулы в специальных карманах, образованных складками полипептидных цепочек глобина. Гемоглобин при нормальном функционировании может находиться в одной из трех форм: феррогемоглобин (обычно называемый дезоксигемоглобином или просто гемоглобином), оксигемоглобин и ферригемоглобин (называемый также метгемоглобином). В феррогемоглобине железо находится в закисной форме Fe(II), одна из двух связей, перпендикулярных к плоскости порфиринового кольца, направлена к атому азота гистидинового остатка, а вторая связь свободна (рис.1, б). Кроме этого гистидинового остатка, называемого проксимальным (соседним), по другую сторону порфиринового кольца и на большем расстоянии от него находится другой гистидиновый остаток — дистальный гистидин, не связанный непосредственно с атомом железа. Взаимодействие молекулярного кислорода со свободным гемом приводит к необратимому окислению атома железа гема [Fe(II) Þ Fе(III); гем Þ гемин]. В дезоксигемоглобине глобин предохраняет железо гема от окисления.
Реакция оксигенации Обратимое присоединение кислорода (оксигенация), позволяющее гемоглобину выполнять свою основную функцию переносчика, обеспечивается возможностью образовать прочные пятую и шестую координационные связи и перенести электрон на кислород не от железа (то есть окислить Fe2+), а от имидазольного кольца проксимального гистидина. Это схематически изображено на рис.1, б. Вместо молекулярного кислорода железо гема может присоединить окись углерода СО (угарный газ). Даже небольшие концентрации СО приводят к нарушению кислородпереносящей функции гемоглобина и отравлению угарным газом.
Выше было сказано, что одна молекула гемоглобина содержит четыре субъединицы и, следовательно четыре гема, каждый из которых может обратимо присоединить одну молекулу кислорода. Поэтому реакцию оксигенации можно разделить на четыре стадии:
Hb+O2 Û HbO2 (1a)
НbO2+O2 Û Hb(O2)2 (1б)
Hb(O2)2+O2 Û Hb(O2)3 (1в)
Hb(O2)3+O2 Û Hb(O2)4 (1г)
Прежде чем рассмотреть эту главную функциональную реакцию гемоглобина более детально, необходимо сказать несколько слов о мышечном гемоглобине — миоглобине. Этот красящий белок поперечнополосатых мышц представляет собой комплекс гема с "четвертушкой" глобина. Он содержит одну молекулу гема и одну полипептидную цепочку, состав и структура которой подобны составу и структуре b-субъединицы гемоглобина. Как и для гемоглобина, важнейшей функцией миоглобина является обратимое присоединение молекулярного кислорода. Эту функцию характеризует так называемая кривая оксигенации, связывающая степень насыщения гемоглобина кислородом (в процентах) с парциальным давлением последнего, рО2 (мм Hg). Типичные кривые оксигенации гемоглобина и миоглобина (при условии достижения химического равновесия) приведены на рис.2, а, б. Для миоглобина кривая является гиперболой, как и должно быть в случае одностадийной химической реакции при условии достижения химического равновесия:
Mb+O2ÛMbO2 (2)
где Mb - миоглобин.
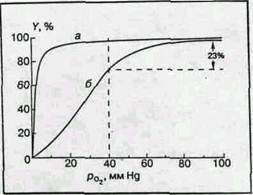
Рис. 2 Кривые оксигенации
миоглобина (а) и гемоглобина (б)
Совершенно другая картина возникает в случае гемоглобина. Кривая диссоциации имеет S-образную форму. Без кислорода молекулы гемоглобина обладают низким сродством к кислороду и равновесие реакции (1а) сдвинуто влево. Затем кривая становится круче и при высоких значениях рО2 практически сливается с кривой диссоциации миоглобина. [2]
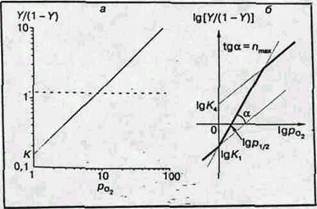
Рис. 3. Логарифмические анаморфозы кривых оксигенации миоглобина (a) и гемоглобина (б)
М.Перутц пишет, что распределение молекул кислорода по молекулам гемоглобина следует библейской притче: "Каждому, у кого есть, дай еще, и у него будет избыток; у того же, у кого нет, забери то немногое, что у него осталось". Это заставляет предположить, что между гемами одной молекулы гемоглобина существует некоторая связь, благодаря которой присоединение кислорода к одному гему влияет на присоединение кислорода к другому гему той же молекулы. Это явление было известно задолго до работ Перутца и установления структуры гемоглобина и механизма его реакции с кислородом. Оно получило название гем-гем взаимодействия. Физиологический смысл гем-гем взаимодействия очевиден. Сигмоидная форма кривой диссоциации создает условия максимальной отдачи кислорода при переносе гемоглобина от легких с высоким значением pO2 к тканям с низким значением pO2. Для человека значения pO2 артериальной и венозной крови в нормальных условиях (Т 37°С, pH 7,4) равны соответственно 100 и 40 ммHg. При этом (рис.2, б) гемоглобин отдает тканям 23% связанного кислорода (степень оксигенации меняется от 98 до 75%). При отсутствии гем-гем взаимодействия для одногемового миоглобина (рис.2, а) эта величина не превышает 5%. Миоглобин поэтому служит не переносчиком, а депо кислорода и отдает его мышечной ткани лишь при резкой гипоксии, когда насыщение ткани кислородом падает до недопустимо низкого значения. [2]
Логарифмическая анаморфоза кривой диссоциации гемоглобина человека представлена на рис.3, б. В этом случае начало кривой представляет собой прямую под углом 45° к координатным осям, как и для миоглобина: первые молекулы кислорода соединяются в основном с молекулами гемоглобина, еще не содержащими кислорода, и гемы таким образом оксигенируются независимо. Это свидетельствуют о том, что гем-гем взаимодействие обусловлено не просто наличием нескольких гемов в молекуле, а тем, что после оксигенации одного гема меняются условия оксигенации других гемов той же молекулы. Затем наклон кривой увеличивается. Тангенс угла максимального наклона получил название коэффициента Хилла (n), который отражает степень кооперативности процесса. Для миоглобина n=1, а для гемоглобина человека (в норме) n~3. Вблизи области полного насыщения гемоглобина кислородом наклон кривой снова становится равным 45° (большинство молекул гемоглобина либо не содержат свободных гемов, либо имеют лишь один гем, способный присоединить кислород).
Механизм кооперативности В 1963 году Моно, Шанже и Джекоб обнаружили, что активность некоторых ферментов меняется скачком между двумя значениями при воздействии на белок некоторых низкомолекулярных агентов, не принимающих участия непосредственно в каталитическом акте. Такие ферменты получили название аллостерических, а само явление – аллостерии. Предполагается, что эти ферменты могут находиться в разных состояниях, переключение между которыми осуществляется при присоединении специфического низкомолекулярного лиганда (необязательно вблизи активного центра). В 1965 году Моно, Уайман и Шанже поняли, что гемоглобин, не являясь ферментом, принадлежит к тому же классу белков. К тому времени уже было известно, что структуры глобул оксигемоглобина и гемоглобина различны, и авторы предположили, что состояния с разными значениями констант оксигенации соответствуют различным пространственным структурам белка. Для гемоглобина постулируется наличие двух таких состояний: R (от англ. relaxed) и Т (от англ. tense). Состояние R характеризуется высоким, а Т – низким сродством к О2 (сильнее и слабее связывают молекулярный кислород соответственно). В рамках этой концепции считается, что как в R-, так и в Т-состоянии сродство к кислороду субъединиц одной глобулы (т.е. есть всех четырех гемов одной глобулы) одинаково. Этот постулат позволяет построить сравнительно простую математическую модель кооперативных свойств гемоглобина: КR, КТ и L (константы равновесия реакций ассоциации в состояниях R, Т и отношение числа молекул гемоглобина в состояниях Т и R соответственно). На рис.2, б ясно, что КТ<КR. Очевидно, увеличение константы ассоциации при переходе из состояния Т в состояние R соответствует в расчете на один гем изменению свободной энергии системы
DG = 2,3 RT lg(KR/KT) кДж/моль. (7)
Для гемоглобина человека при 37°С эта "свободная энергия кооперативности" равна 5,61 кДж/моль. В физиологических условиях при отсутствии кислорода лишь ~ 3·10-5 % молекул гемоглобина находятся в R-форме, а в условиях полного насыщения кислородом лишь ~ 7-10-3 % — в Т-форме. [2]
Изменения электронной и пространственной структуры гемоглобина в процессе оксигенации На рис.4 схематически показаны электронная структура железа гема, положение атома железа относительно плоскости порфиринового кольца гема, спектральные и магнитные характеристики молекул в различных состояниях молекулы гемоглобина: дезоксигемоглобин, оксигемоглобин и ферригемоглобин. Следует подчеркнуть, что во всех случаях речь идет о равновесных состояниях молекул белка. Мы увидим далее, что переход из одного состояния в другое требует значительного (в молекулярных масштабах) времени, в течение которого система проходит через несколько неравновесных состояний, заметно отличающихся по своим физическим и химическим свойствам от равновесных. [2]
В молекуле дезоксигемоглобина железо отстоит от плоскости порфиринового кольца примерно на 0,5-0,6 А° (есть небольшие отличия между a- и b-субъединицами). Из шести 3d электронов железа Fe(II) два электрона спарены на одной из низших d-орбиталей (dxy, dyz, dxs), а четыре электрона занимают оставшиеся d-орбитали, их спиновые моменты, согласно правилу Хунда, параллельны и суммарный спин S-2. Магнитный момент гема в этом состоянии равен ~ 5,5 боровского магнетона (БМ), а спектр поглощения в зеленой области имеет характерную полосу с lmax ~ 556 нм. Присоединение кислорода ведет к значительным изменениям. Атом железа в оксигемоглобине лежит практически в плоскости порфиринового кольца (расстояние до плоскости составляет 0,16 А° в α- и 0,00 А° в β- субъединицах). Все шесть d-электронов спарены на трех низших d-орбиталях, S= 0, оксигемоглобин диамагнитен. В зеленой области спектра имеются две характерные полосы поглощения: а (lmax 576 А°) и b (542 А°).
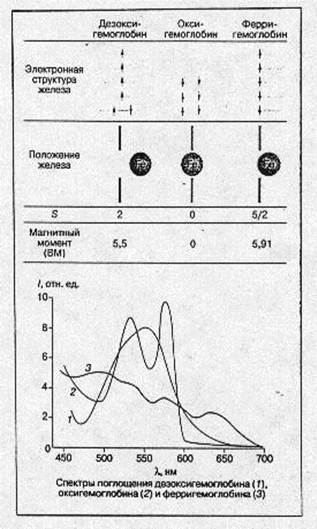
Рис.4. Основные характеристики молекулы гемоглобина в различных состояниях.
В ферригемоглобине (метгемоглобин) при нейтральных значениях рН место кислорода занимает молекула воды (при щелочных значениях рН-ОН), железо находится значительно ближе к плоскости гема, чем в дезоксигемоглобине, все пять d-электронов неспарены и занимают пять d-орбиталей. S = 5/2 и магнитный момент равен 5,91 БМ.
Структурные изменения в активном центре (вблизи гема) приводят и к значительным изменениям пространственной структуры всего белка. При оксигенации (переход от Т- к R-форме) смещение отдельных аминокислотных остатков достигает 7 А°. Как уже было сказано выше, четвертичная структура гемоглобина характеризуется наличием четырех полипептидных цепей, образующих две (a- и две b-субъединицы). Более детальные исследования показали, что субъединицы образуют ab-димеры. ТÞR – переход сопровождается поворотом одного димера относительно другого на 12-15° и в конечном счете приводит к увеличению карманов, в которых находятся гемы. Эти структурные изменения инициируются присоединением первой молекулы О2 к одному из свободных гемов и распространяются на всю глобулу. Именно поэтому в равновесной смеси всегда присутствуют только Т- и R-формы. Эти димеры в Т-форме стягиваются 14 дополнительными (по сравнению с R-формой) солевыми мостиками (водородные связи между ионными или нейтральными группами аминоксилот, ван-дер-ваальсовы контакты). Кроме того, между b-субъединицами в Т-форме присоединяется молекула дифосфоглицерата, что также приводит к сужению карманов. Эти изменения схематически представлены на рис.5.
Триггером для всех описанных выше структурных перестроек при переходах между Т- и R-формами и обратно служит присоединение или отщепление кислорода. После локального элементарного химического акта: присоединение или отщепление низкомолекулярного лиганда, окисление железа при образовании ферригемоглобина (иначе говоря, после появлении лишнего положительного заряда на железе) – возникает существенно неравновесное конформационное состояние – изменения вблизи активного центра уже произошли, а вся огромная молекула белка осталась в прежнем, еще не отрелаксировавшем состоянии. Последующая релаксация может занимать микросекунды и даже секунды. В ходе этой релаксации меняются не только физические, но и химические свойства белка, в частности скорости последующих химических актов, если они успевают произойти до полного завершения релаксации. Таким образом, описанная выше картина процессов, сопровождающих обратимое связывание кислорода гемоглобином, является лишь первым, хотя и очень важным приближением к истине. Так, например, быстрое восстановление железа в ферригемоглобине коротким (микро- или наносекунды) импульсом электронов приводит к возникновению неравновесного состояния, в котором железо уже восстановлено, но не отошло от плоскости порфиринового кольца. По спектральным и магнитным характеристикам это состояние соответствует равновесному оксигемоглобину. Релаксация гема и его ближайшего окружения с удалением железа от плоскости порфиринового кольца занимает при комнатной температуре десятки микросекунд, а полная релаксация всей белковой глобулы к равновесной Т-форме дезоксигемоглобина — сотни миллисекунд.
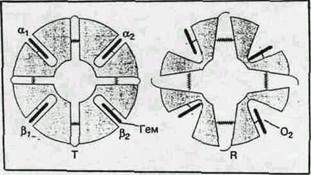
Рис. 5. Структурная схема перехода гемоглобина от Т- к R-форме
Другие реакции и функции гемоглобина При взаимодействии молекулярного кислорода с гемоглобином существует небольшая, но конечная вероятность окисления последнего: молекула О2 не присоединится, но окислит железо: Fe2+ + O2 Þ Fe3+ O2-. Поэтому при дыхании в эритроцитах непрерывно образуется метгемоглобин. Для его восстановления в эритроците существует специальная ферментативная система, восстанавливающая метгемоглобин и превращающая его в нормальный дезоксигемоглобин. При нарушении этой системы возникает тяжелое заболевание — метгемоглобинемия, при котором гемоглобин перестает быть переносчиком кислорода.
Гены, ответственные за синтез гемоглобина, могут подвергаться мутациям, меняющим структуру и функции белка. Наиболее изучена мутация, приводящая к замене только одной аминокислоты в полпептидных цепочках b-субъединиц гемоглобина. Замена глутамина на валин ведет к тяжелой болезни – серповидноклеточной анемии: эритроциты принимают форму серпа и теряют способность переносить кислород.
Присоединение кислорода меняет кислотно-основные свойства гемоглобина. Оксигемоглобин является более сильной кислотой, чем дезоксигемоглобин. Поэтому в тканях, где значительная часть гемоглобина теряет кислород и становится более сильным основанием, гемоглобин связывает образующуюся в ходе метаболических внутриклеточных процессов углекислоту. В альвеолах легких дезоксигемоглобин снова превращается в оксигемоглобин, становится более сильной кислотой и способствует отщеплению СО2. Это слегка упрощенное описание важного процесса транспорта углекислоты эритроцитами. Углекислота, освобождаемая тканями, недостаточно хорошо растворима для эффективного переноса. С помощью фермента карбоангидразы, ускоряющего прямую и обратную реакцию
СО2+Н2ОÛНСО3-+Н+ (8)
двуокись углерода превращается в хорошо растворимый бикарбонат-анион. В капиллярах тканей отщепление кислорода повышает содержание дезоксигемоглобина, связывающего протоны и смещающего, таким образом, равновесие реакции (8) вправо. Легко растворимый ион бикарбоната переносится кровью. В альвеолах легких гемоглобин оксигенируется, протоны освобождаются и равновесие (8) смещается влево. Образуется плохо растворимая двуокись углерода СО2, которая удаляется из водной фазы и выдыхается. Таким образом, гемоглобин работает как буфер с переменным значением pK. Функция гемоглобина как переносчика углекислоты не менее важна, чем его функция переносчика кислорода.
Гемоглобин – одно из наиболее хорошо изученных белков. Десятки лет исследований гемоглобина в описании и понимании физических, химических и биологических аспектов его функционирования. Огромный вклад внесли работы Макса Перутца и его сотрудников в Кавендишской лаборатории (Кембридж, Великобритания). Однако важность этих работ касается не только гемоглобина. Они послужили основой развития современных представлений о механизмах ферментативного катализа, связав непосредственно кинетику и термодинамику биохимических реакций с динамикой конформационных изменений макромолекул белка. Если отвлечься от непосредственной практической пользы полученных результатов для медицины, фармакологии, то фундаментальное значение работ по изучению механизма функционирования гемоглобина заключается в стимулировании прогресса в установлении законов протекания важнейших процессов: ферментативного катализа и внутриклеточной трансформации энергии в биологических системах. [2]
Об участии гемоглобина в мембранной организации эритроцитов свидетельствует феномен образования серповидной и других форм эритроцитов при различных гемоглобинопатиях, первооснову которого и составляет аномальное взаимодействие эритроцитарной мембраны с мутантными гемоглобинами. Не исключено, что изменение упруго-механических свойств эритроцитарных мемран при повышении отрицательного заряда их внешней поверхности происходит вследствие улучшения условий для создания и структурирования белкового слоя, формируемого с участием гемоглобина на внутренней поверхности эритроцитарных мембран. [13]
Показано, что при получении безгемоглобиновых теней эритроцитов в препаратах электронно-микроскопически обнаруживается большое количество везикул диаметром примерно 100 нм, что свидетельствует о частичной фрагментации плазматической мембраны эритроцитов. В то же время если в мембранах содержится много гемоглобина и других белков, обнаруживаемых в примембранных слоях, то фрагментации мембраны определяемой таким образом, не наблюдается. Имеются и другие доказательства стабилизирующего действия гемоглобина на эритроцитарные мембраны. Например, S.Knutton и соавторам удалось получить две фракции эритроцитарных мембран, различающихся содержанием связанного с ними гемоглобина. Ими было показано, что во фракции эритроцитарных мембран с большим содержанием гемоглобина разрушение липопротеиновой структуры мембран при их дегидротации происходит намного слабее, чем в случае с фракцией эритроцитарных мембран с малым содержанием гемоглобина. В основе изменения условий для структурирования белково-образующего примембранного слоя внутри эритроцитов может лежать изменение электрических поверхностных свойств на внешней стороне. Это возможно вследствие того, что для мембран в липидной зоне дебаевский радиус экранировки зарядов гораздо больше ее толщины, в результате поверхностные заряды с обеих сторон эритроцитарных мембран взаимно влияют друг на друга, образуя самосогласованную систему. Кроме того, условия для образования белкового слоя, стабилизирующего мембраны эритроцитов, могут изменяться и в результате защелачивания их внутренней среды, что имеет место при суспендировании эритроцитов в щелочной или неэлектролитной среде, например, сахарозной. Это может способствовать гемоглобину и другим примембранным белкам за счет изменения их свойств образовывать прочный примембранный белковый слой (в отношении гемоглобина, например, известно, что его свойства, в том числе кислородосвязывающие, сильно изменяются при варьировании рН). Вместе с тем защелачивание внутренней среды эритроцитов не всегда может служить фактором, стабилизирующим их структуру. В литературе, например, утверждают, что для эритроцитов новорожденного теленка, для которых характерно наличие фетального гемоглобина, любые экспериментальные манипуляции, результатом которых является внутриклеточное защелачивание среды, вызывают их самопроизвольный гемолиз. По мере развития организма теленка (2-3 месяца жизни) эритроциты с нормальным гемоглобином, появляющиеся взамен вытесняемых из кровеносного русла клеток, содержащих фетальный гемоглобин, становятся устойчивыми к внутриклеточному защелачиванию их среды. При этом важно отметить, что, по данным литературы, в гемолизе фетальных эритроцитов при защелачивании их внутренней среды, помимо фетального гемоглобина, определенная роль принадлежит и мембранному белку band III. [13]
Приведенные литературные данные дают основание сделать вывод о тесной взаимосвязи процессов метаболизма, трансмембранного транспорта, изменений формы и механических свойств эритроцита, что, по-видимому, позволяет организму осуществлять координированную регуляцию функционирования клетки через соответствующие рецепторные структуры, имеющиеся на наружной поверхности мембраны. [9]
Таким образом, в поддержании структурной целостности эритроцитов важное значение имеют внутренние примембранные белковые слои, структура и взаимодействие которых с эритроцитарными мембранами взаимно обусловлены и в целом представляют собой единую структурную организацию. Поэтому всякая модификация как самой мембраны, так и содержащегося в них гемоглобина в конечном счете сопровождается и модификацией этой особой организации, частным примером проявления которой может быть изменение ее механических свойств. [13] Приведенные данные побудили нас провести исследование с целью оценки количественного содержания мембрансвязанного гемоглобина эритроцитов человека и его связи со структурными белками.
материал и методы исследования
Материал исследования
Материалом настоящего исследования послужили эритроциты 124 практически здоровых лиц, проживающих в городе Курске, в возрасте от 18 до 47 лет. Набор добровольцев проводился при исключении любой соматопатологии.
У обследуемых проводился забор крови из локтевой вены в сухую стерильную посуду в количестве 5-7 мл с целью получения образов эритроцитарных мембран.
Методы исследования
Для достижения поставленной цели и решения задач настоящей работы использовался комплекс методов.
Биохимические методы.
Реактивы и биоматериалы. В работе использовались: декстран Т-500 фирмы "SIGMA" (США), HBS – целлюлоза фирмы "SIGMA" (США), гидрофосфат натрия, хлористый натрий, мочевина фирмы "Bio-Rad" (США), трис, 2 – меркаптоэтанол, персульфат аммония "Reanal" (Венгрия), додецилсульфат натрия (ДДС) фирмы "Диа-Фарм" (Россия), акриламид "SIGMA" (США), N,N¢ - метилен бисакриламид "FLUKA" (Швейцария), кумасси G – 250 фирмы "Serva" (ФРГ), бромфеноловый синий, ТЕМЕД, глицин, набор белков для определения молекулярного веса MS-2 (Россия).
Реактивы отечественного производства были марки "ХЧ" и "ОСЧ".
Получение эритроцитов. Эритроциты получали из 5 мл гепаринизированной крови по методу Бейтлера с незначительной модификацией. [14] Сначала гепаринизированную кровь отстаивали дважды в растворе PBS, содержащем 3% декстран Т –500, в течение 30 минут. Затем эритроцитарную массу подвергали дополнительной очистке, пропуская ее через колонку с HBS-целлюлозой, после чего эритроциты ценрифугировали при 3000 оборотах в минуту. Далее из полученной очищенной массы эритроцитов проводили выделение мембран.
Выделение мембран. Для получения препаратов мембран эритроциты разрушали осмотическим шоком по методу Dodge с небольшой модификацией, заключавшейся в том, что гемолиз эритроцитов и отмывку "теней" проводили двукратно в 10 мМ Na – фосфатном буфере с добавлением ингибитора протеназ PMSF. После этого мембраны эритроцитов лиофилизировали и хранили при -20°С.
Одномерный электрофорез по Лэммли. Электрофорез в присутствии ДДС проводили модифицированным методом методом Лэммли. [14] Для этой цели использовали следующие растворы:
1. 60% акриламид –0,8% метилен бисакриламид.
2. Буфер для разделяющего геля: 1 М трис – HCI (pH 8,8).
3. Буфер для концентрирующего геля: 0,5 М трис – HCI (pH 6,8).
4. 10% додецилсульфат натрия.
5. 10% персульфат аммония.
6. ТЕМЕД.
7. Электродный буфер для ЭФ: 0,025М трис, 0,193М глицин, 0,1% додецилсульфат натрия.
Растворы хранились при +4°С две-три недели. Раствор 5 готовился перед использованием.
Электрофорез проводили в пластинах ПААГ размером 180х180х1мм, приготовленных с линейным градиентом концентрации акриламида 5 - 25%.
Для приготовления одной пластины ПААГ брали: 0,85 мл раствора 1, 5,5 мл дистиллированной воды, 3,6 мл раствора 2, 101 мкл раствора 4, 5 мкл раствора 6 и 20 мкл раствора 5 (легкий раствор, 5%). В тяжелом (25%) растворе в отличие от легкого бралось 4,2 мл раствора 1 и 2,6 мл дистиллированной воды. Через смеситель эти растворы подавались перистальтическим насосом в формирователь пластины, в течение 10 минут заполняя пластину на 4 см ниже верхнего края. Сверху на полимеризующуюся смесь наслаивалось 0,5 мл дистиллированной воды. Так формировался разделяющий гель. Полимеризация продолжалась в течение 30-40 минут. После полимеризации воду удаляли и заливали раствор, формирующий концентрирующий гель. Для приготовления такого раствора брали 660 мкл раствора 1, 100 мкл раствора 4, 2,5 мл раствора 3, 4 мкл раствора 6, 70 мкл раствора 5 и 6 мл дистиллированной воды. Вставляли формирователь лунок и наслаивали 0,3 мл дистиллированной воды. Полимеризация продолжалась в течение 20 минут, после чего пластины ПААГ могли храниться при =4°С в течение двух суток.
Подготовка проб Для приготовления анализируемого образца для ЭФ брали 1 мг лиофилизированных мембран эритроцитов и растворяли в 40 мкл уравновешивающего буфера, в состав которого входили:
(на 10 мл уравновешивающего буфера)
1. 3 г мочевины;
2. 0,2 г ДДС;
3. 1,25 мл раствора 3;
4. 0,5 мл меркаптоэтанола;
5. 5 мкл красителя бромфенолового синего.
Уравновешивающий буфер хранили при температуре -20°С в течение месяца.
Подготовленные таким образом анализируемые образцы стояли в течение одного часа при комнатной температуре, после чего их вносили объемом 15 мкл в сформированные в концентрирующем геле лунки.
Электрофорез проводили при силе тока 35 мА, пока напряжение не возрастало до 300 В, затем стабилизировали источник питания по данному напряжению и проводили ЭФ этом режиме, пока лидирующий краситель не доходил 1 см до края пластины.
После ЭФ фореграммы окрашивали в течение одного часа красителем Кумасси G - 255 по модифицированной методике Fairbanks в растворе, содержащем 10% уксусной кислоты, 25% изопропанола, 0,05% кумасси голубого. После этого несвязавшуюся краску отмывали в течение 12 часов 10%-ной уксусной кислотой до полного исчезновения фонового окрашивания.
Отмытые фореграммы затем дегидратировали в течение 30 минут в растворе, содержащем 280 мл изопропанола, 25 мл глицерина и 195 мл дистиллированной воды. Далее пластины ПААГ плотно фиксировали между двумя слоями целлюлозной бумаги и в натянутом виде высушивали при комнатной температуре.
На электрофореграммах идентифицировали 17 белковых фракций.
Денситометрирование. Денситометрирование электрофореграмм проводили на лазерном денситометре «Ultrascan XL». Молекулярную массу белков определяли с помощью маркерных белков с известной молекулярной массой: бычий сывороточный альбумин (Mr=68 kD), овальбумин (Mr=43 kD), химотрипсиноген (Мr=25 kD), миоглобин (Мr=17,5 kD) и цитохром (Мr=12,4 kD).
Определение концентрации белка. Содержание белковых фракций в исследуемом образце определяли по известной массе маркерного белка бычьего сывороточного альбумина, через полученные при деситометрировании площади альбумина и каждой белковой фракции в отдельности по формуле:
m(x)=S(x)´4.5/S(a),
где m(x) – масса белковой фракции,
S(x) – площадь этой фракции под пиком на денситограмме
4.5 – масса маркерного белка альбумина в мкг,
S(a) – площадь маркерного белка альбумина под пиком на денситограмме.
Дальнейший пересчет количественного содержания белковых фракций проводили на 1 миллиграмм общего белка.
Методы статистической обработки
Статистическая обработка материала проведена на ПВМ IBM PC/AT (486) с использованием программы "Gen 1", составленной д.б.н Трубниковым В.И. и пакета прикладных программ "STATGRAPHICS v3.0".
При описании количественных признаков использовались параметры нормального распределения: среднее значение, стандартная ошибка среднего значения, несмещенная дисперсия. Для проверки статистических гипотез использовались параметрические критерии Стьюдента и Фишера. Уровень значимости принимали равный 0,05.
Кластерный анализ. При проведении кластерного анализа в качестве объектов выступали индивиды и изучаемые признаки, а мерой сходства служили коэффициенты корреляции без учета знака. Результаты кластерного анализа позволили определить соответственно некоррелированные группы изучаемых параметров, детерминирующих фенотипическую изменчивость признаков. На каждом шаге кластеризации выбирался наибольший по значению элемент матрицы, стоящий на пересечении i-й строки и j-го столбца объединения i-го и j-го признаков в кластер, рассматриваемый как новый признак. Для выделения кластера пересчитывались значения коэффициентов корреляции в матрице с тем, чтобы на следующем шаге кластеризации поиск максимального коэффициента корреляции проводился с учетом предыдущих результатов. Приведенный алгоритм кластерного анализа служил лишь для подразделения корреляционной матрицы на отдельные подсистемы без проверки статистической значимости уровней объединения кластеров. Проверка гипотезы о некоррелированности выделенных подсистем осуществлялась с использованием специальных критериев.
РЕЗУЛЬТАТЫ СОБСТВЕННЫХ ИССЛЕДОВАНИЙ
Количественное содержание гемоглобина и основных белков мембран эритроцитов человека.
В ходе данной работы нами было проведено изучение количественного содержания основных белков эритроцитарных мембран и мембрансвязанного гемоглобина. Полученные результаты представлены в таблице 1.
В зависимости от электрофоретической подвижности белки были разделены на 15 фракций, 16-ю составил гемоглобин. При электрофорезе белковые фракции расположились по мере уменьшения молекулярных масс белков. Из таблицы 1 видно, что наиболее представительными белками в мембранах эритроцитов человека являются анионтранспортный белок полосы 3, a- и b-спектрины и актин. Второе место по количественному содержанию заняли белки полосы 4.5 (транспортер глюкозы и нуклеозидов), белки полосы 7 (основным белком этой полосы является тропомиозин) и белок полосы 4.2 (паллидин).
Менее представительными оказались анкирины 2.1, 2.2, и 2.3, белок полосы 4.1, глицеральдегид-3-фосфатдегидрогеназа и глутатион-S-трансфераза. Наименьшим содержанием отличался белок полосы 4.9 и мембрансвязанный гемоглобин. Количественное содержание гемоглобина составило 17,40±0,74 мкг на 1 мг общего белка мембран. Кроме того, гемоглобину соответствовало и наименьшее значение дисперсии (d2= 66,91).
Проведенное исследование количественного содержания основных белков мембран эритроцитов позволило оценить их представительность. Полученные нами показатели согласуются с литературными данными. [4, 5]
Взаимное варьирование количественной представительности отдельных белков эритроцитов человека.
С целью установления особенностей взаимного варьирования содержания белковых фракций в анализируемых образцах нами был проведен многомерный количественного содержания белков эритроцитарных мембран человека. Была построена матрица фенотипических корреляций количественного содержания гемоглобина и основных белков мембран эритроцитов. Данная матрица представлена в таблице 2. Из нее видно, что полученные коэффициенты корреляции имели как положительную, так и отрицательную направленность.
Для установления приоритетности во взаимосвязях вариабельности количественного содержания гемоглобина и основных мембранных белков эритроцитов нами был проведен кластерный анализ. Полученные результаты представлены на рисунке 6.
Кластерный анализ корреляционной матрицы позволил выделить 4 группы взаимнокоррелирующих белков по их количественному содержанию. Первую группу составили a- и b-спектрины, белки полосы 4.1, 4.2 (r=0,255). В состав второй группы вошли анкирины 2.1, 2.2 и 2.3 (r=0,278). Третью группу образовали гемоглобин, тропомиозин, глютатион-S-трансфераза и белок полосы 4.5.1 (r=0,316). Четвертую группу составили актин, белки полосы 4.5, глицеральдегид-3-фосфатдегидрогеназа и анионтранспортный белок полосы 3 (r=0,209). Исключение составила фракция 4.9, вариабельность количественного содержания которой носила независимый характер.
Низкий уровень объединения наблюдался между количественным содержанием анкирином 2.3 и анкирином 2.1 и 2.2 (r=0,278), актином и белком полосы 4.5 (r=0,327).
Рисунок 6
Дендрограмма фенотипических корреляций количественного содержания гемоглобина и основных белков мембран эритроцитов человека
N = 124 R (5%) = 0,176
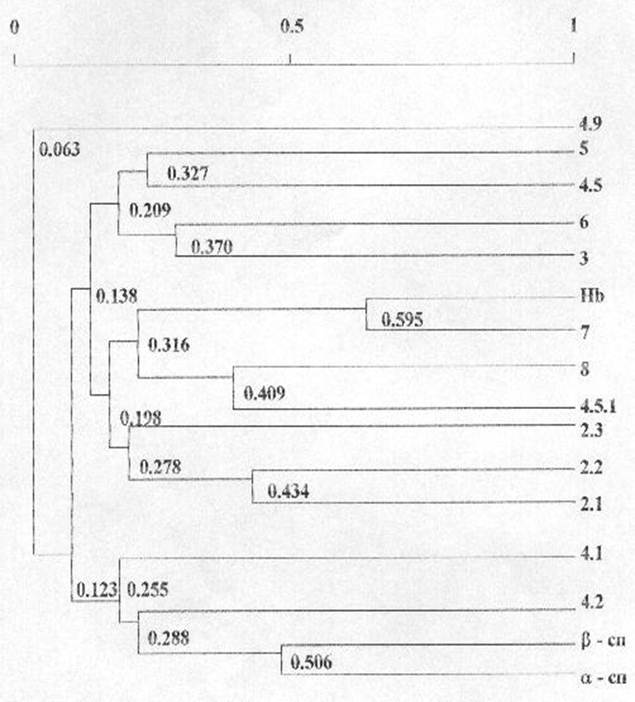
Группу кластеров со средним уровнем
объединения составили анкирины 2.1 и 2.2 (r=0,434),
глутатион-S-трансфераза и белок полосы 4.5.1 (r=0,409),
глицеральдегид-3-фосфатдегидрогеназа и белок полосы 3 (r=0,370).
Высокий уровень объединения имел место между количественным содержанием a- и b-спектринов (r=0,506)
Самый высокий уровень объединения (r=0,595) соответствовал количественному содержанию гемоглобина и белков полосы 7, основным составляющим белком которой является тропомиозин.
Обсуждение
На основе полученных данных установлено, что наиболее представительными белками в мембранах эритроцитов человека являются анионтранспортный белок полосы 3, a- и b-спектрины, а также актин. Наименьшее количественное содержание соответствует мембрансвязанному гемоглобину. Несмотря на то, что он содержится в мембране эритроцитов человека в меньшем количестве, гемоглобин играет важную роль как в образовании и поддержании стабильности цитоскелета, так и в механизмах ферментативного катализа и внутриклеточной трансформации энергии.
Полученные данные позволили предположить, что в эритроцитарных мембранах рассматриваемые белки во взаимном варьировании их количественного содержания образуют 4 группы взаимнокоррелирующих систем. Причем каждая из этих групп по отношению к остальным характеризуется относительной независимостью.
Согласно результатам нашего исследования количественное содержание гемоглобина наиболее тесно связано с содержанием тропомиозина. О причинах этого пока судить трудно вследствие недостаточного количества данных, касающихся этого вопроса - в доступной нам литературе он не получил должного освещения. Но мы надеемся, что полученные данные послужат основой для дальнейшей детализации в изучении связи гемоглобина со структурными белками.
Таким образом, полученные нами данные свидетельствуют о сложной структурной и функциональной взаимосвязи белков в мембранах эритроцитов.
заключениеЦитоплазматические мембраны эритоцитов человека играют ключевую роль в обеспечении и регуляции физиологической активности этих клеток. Специфические функции мембран обеспечивает сложная структурная организация, главным компонентом которой являются белки.
Современные биохимические и биофизические методы позволили выделить практически все основные мембранные белки эритроцитов, изучить их биохимическую структуру и основные аспекты функционирования. Вместе с тем в научной литературе не получил должного освещения вопрос о структуре мембрансвязанного гемоглобина, о его связи со структурными белками. Нами была проведена оценка количественного содержания гемоглобина и основных белков мембран эритроцитов человека, так как нормальное функционирование мембраны определяется не только присутствием белковых компонент, но и их количественным содержанием.
Кроме того был предпринят анализ взаимоварьирования количественного содержания гемоглобина и структурных белков. В ходе исследования нами был установлен факт взаимозависимости количественного содержания гемоглобина и тропомиозина. Результаты данного исследования могут служить основой для дальнейшей детализации в изучении связи гемоглобина со структурными белками. Нами планируется провести ряд исследований с целью изучения количественного содержания гемоглобина и его взаимосвязи со структурными белками мембран эритроцитов не только в норме, но и при различных патологических состояниях. С тем, чтобы прогнозировать наряду с уровнем данных нарушений их связь между собой и влияние на них изменений окружающей среды.
выводы
1. Количественное содержание гемоглобина в мембранах эритроцитов составляет 17,40±0,74 мкг на 1 мг общего белка мембран и является наименьшим, по сравнению с количественным содержанием остальных мембранных белков.
2. Вариабельность количественного содержания гемоглобина характеризуется достаточно высокой сопряженностью с количе37ственным содержанием белка полосы 7 – тропомиозина.
Литература
1. Балашов В.И., Резаев А.А., Ярыга В.В. Содержание гемоглобина и его дериватов в крови детей, проживающих в населенных пунктах санитарно-защитной зоны астраханского газового комплекса// Педиатрия. – 1995. - №2. – С.75-77.
2. Блюменфельд Л.А. Гемоглобин// Соросовский образовательный журнал. – 1998. - №4. – С.33-38.
3. Гааль Э., Медьеши Г., Верецки Л. Электрофорез в разделении биохимических макромолекул. – М. – 1982. – 446с.
4. Гончаренко М.С., Андрух Г.А., Рязанцев В.В. Белковый спектр эритроцитарных мембран в норме и при псориазе// Вестник дерматологии и венерологии. – 1989. - №3. – С.4-7.
5. Гончарова Е.И., Пинаев Г.П. Белки цитоскелета эритроцитов// Цитология. – 1988. – т.30, №1. – С.5-18.
6. Громов П.С., Захаров С.Ф., Шишин С.С., Ильинский Р.В. Двумерная карта мембранных белков эритроцитов человека// Биохимия. – 1988. – т.53, вып.8. – С.1316-1326.
7. Громов П.С., Шандала А.М., Ковалев Л.И., Шишкин С.С. Изучение белков мембран эритроцитов человека методом двумерного электрофореза// Бюллетень экспериментальной биологии и медицины. – 1986. - №7. – С.28-30.
8. Казеннов А.М., Маслова М.Н. Влияние мембранного скелета безъядерных эритроцитов на свойства транспортных АТФаз// Цитология. – 1991. – т.33, №11. – С.32-41.
9. Казеннов А.М., Маслова М.Н. Структурно-биохимические свойства мембраны безъядерных эритроцитов// Физиологический журнал СССР им. И.М.Сеченова. – 1987. – т.73, №12. – С.1587-1594.
10. Казеннов А.М., Маслова М.Н., Шагабодов А.Д. Роль белков мембранного скелета безъядерных эритроцитов в функционировании мембранных ферментов// Докл. АН СССР – 1990. – т.312. - №1. – С.223-226.
11. Лакин Г.Ф. Биометрия: Учебное пособие для биол. спец. Вузов – 4-е изд., перераб. и доп. – М.: Высш.шк., 1990. – 352с.
12. Остерман Л.А. Методы исследования белков и нуклеиновых кислот. Электрофорез и ультрацентрифугирование. – М.: Наука, 1981. – 288с.
13. Петренко Ю.М., Владимиров Ю.А. Роль поверхностных зарядов в поддержании осмотической резистентности эритроцитов// Гематология и трансфузиология. – 1987. - №10. – С.15-18.
14. Солодилова М.А. Роль генетических и средовых факторов в детерминации количественного содержания основных белков мембран эритроцитов человека/ Дис. на соискание ученой степени к.б.н. – М., 1999. – 160с.
15. Шандала А.М., Захаров С.Ф., Громов П.С., Шишкин С.С. Белковый состав мембран эритроцитов человека, фракционных в ступенчатом градиенте декстрана// Гематология и трансфузиология. – 1987. - №10. – С.28-31.